Мозг, общение нейронов и энергетическая эффективность
По всей видимости, в эволюции сформировались энергетически эффективные механизмы кодирования и передачи информации в мозге. Подпись: «Усердно пытаюсь минимизировать энергозатраты».
Автор
Редакторы
Статья на конкурс «био/мол/текст»: Клеточные процессы, обеспечивающие обмен информацией между нейронами, требуют много энергии. Высокое энергопотребление способствовало в ходе эволюции отбору наиболее эффективных механизмов кодирования и передачи информации. В этой статье вы узнаете о теоретическом подходе к изучению энергетики мозга, о его роли в исследованиях патологий, о том, какие нейроны более продвинуты, почему синапсам иногда выгодно не «срабатывать», а также, как они отбирают только нужную нейрону информацию.

Конкурс «био/мол/текст»-2017
Эта работа опубликована в номинации «Свободная тема» конкурса «био/мол/текст»-2017.

Генеральный спонсор конкурса — компания «Диаэм»: крупнейший поставщик оборудования, реагентов и расходных материалов для биологических исследований и производств.

Спонсором приза зрительских симпатий и партнером номинации «Биомедицина сегодня и завтра» выступила фирма «Инвитро».

Происхождение подхода
С середины ХХ века известно, что головной мозг потребляет значительную часть энергоресурсов всего организма: четверть всей глюкозы и ⅕ всего кислорода в случае высшего примата [1–5]. Это вдохновило Уильяма Леви и Роберта Бакстера из Массачусетского технологического института (США) на проведение теоретического анализа энергетической эффективности кодирования информации в биологических нейронных сетях (рис. 1) [6]. В основе исследования лежит следующая гипотеза. Поскольку энергопотребление мозга велико, ему выгодно иметь такие нейроны, которые работают наиболее эффективно — передают только полезную информацию и затрачивают при этом минимум энергии.
Это предположение оказалось справедливым: на простой модели нейронной сети авторы воспроизвели экспериментально измеренные значения некоторых параметров [6]. В частности, рассчитанная ими оптимальная частота генерации импульсов варьирует от 6 до 43 имп./с — почти так же, как и у нейронов основания гиппокампа. Их можно подразделить на две группы по частоте импульсации: медленные (
40 имп./с). При этом первая группа значительно превосходит по численности вторую [7]. Аналогичная картина наблюдается и в коре больших полушарий: медленных пирамидальных нейронов (
4—9 имп./с) в несколько раз больше, чем быстрых ингибиторных интернейронов (>100 имп./с) [8], [9]. Так, видимо, мозг «предпочитает» использовать поменьше быстрых и энергозатратных нейронов, чтобы те не израсходовали все ресурсы [6], [9–11].
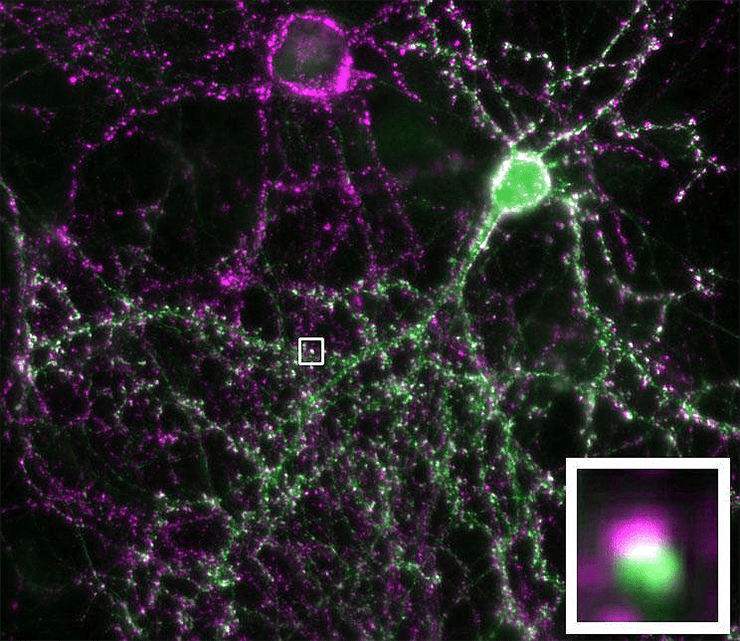
Рисунок 1. Представлены два нейрона. В одном из них фиолетовым цветом окрашен пресинаптический белок синаптофизин. Другой нейрон полностью окрашен зеленым флуоресцентным белком. Мелкие светлые крапинки — синаптические контакты между нейронами [12]. Во вставке одна «крапинка» представлена ближе.
Группы нейронов, связанных между собой синапсами, называются нейронными сетями [13], [14]. Например, в коре больших полушарий пирамидальные нейроны и интернейроны образуют обширные сети. Слаженная «концертная» работа этих клеток обусловливает наши высшие когнитивные и другие способности. Аналогичные сети, только из других типов нейронов, распределены по всему мозгу, определенным образом связаны между собой и организуют работу всего органа.
Что такое интернейроны?
Нейроны центральной нервной системы разделяются на активирующие (образуют активирующие синапсы) и тормозящие (образуют тормозящие синапсы). Последние в значительной степени представлены интернейронами, или промежуточными нейронами. В коре больших полушарий и гиппокампе они ответственны за формирование гамма-ритмов мозга [15], которые обеспечивают слаженную, синхронную работу других нейронов. Это крайне важно для моторных функций, восприятия сенсорной информации, формирования памяти [9], [11].
Интернейроны отличаются способностью генерировать значительно более высокочастотные сигналы, чем другие нейроны. Они также содержат больше митохондрий, главных органелл энергетического метаболизма, «фабрик» по производству АТФ. Последние к тому же содержат большое количество белков цитохром-с оксидазы и цитохрома-с, являющихся ключевыми для метаболизма. Так, интернейроны являются крайне важными и, в то же время, энергозатратными клетками [8], [9], [11], [16].
Работа Леви и Бакстера [6] развивает концепцию «экономии импульсов» Горация Барлоу из Университета Калифорнии (США), который, кстати, является потомком Чарльза Дарвина [17]. Согласно ей, при развитии организма нейроны стремятся работать только с наиболее полезной информацией, фильтруя «лишние» импульсы, ненужную и избыточную информацию. Однако эта концепция не дает удовлетворительных результатов, так как не учитывает метаболические затраты, связанные с нейрональной активностью [6]. Расширенный подход Леви и Бакстера, в котором внимание уделено обоим факторам, оказался более плодотворным [6], [18–20]. И энергозатраты нейронов, и потребность в кодировании только полезной информации являются важными факторами, направляющими эволюцию мозга [6], [21–24]. Поэтому, чтобы лучше разобраться в том, как устроен мозг, стоит рассматривать обе эти характеристики: сколько нейрон передает полезной информации и сколько энергии при этом тратит.
За последнее время этот подход нашел множество подтверждений [10], [22], [24–26]. Он позволил по-новому взглянуть на устройство мозга на самых разных уровнях организации — от молекулярно-биофизического [20], [26] до органного [23]. Он помогает понять, каковы компромиссы между выполняемой функцией нейрона и ее энергетической ценой и в какой степени они выражены.
Как же работает этот подход?
Положим, у нас есть модель нейрона, описывающая его электрофизиологические свойства: потенциал действия (ПД) и постсинаптические потенциалы (ПСП) (об этих терминах — ниже). Мы хотим понять, эффективно ли он работает, не тратит ли неоправданно много энергии. Для этого нужно вычислить значения параметров модели (например, плотность каналов в мембране, скорость их открывания и закрывания), при которых: (а) достигается максимум отношения полезной информации к энергозатратам и в то же время (б) сохраняются реалистичные характеристики передаваемых сигналов [6], [19].
Поиск оптимума
Эти «оптимальные» значения параметров затем нужно сравнить с измеренными экспериментально и определить, насколько они отличаются. Общая картина отличий укажет на степень оптимизации данного нейрона в целом: насколько реальные, измеренные экспериментально, значения параметров совпадают с рассчитанными. Чем слабее выражены отличия, тем нейрон более близок к оптимуму и работает энергетически более эффективно, оптимально. С другой стороны, сопоставление конкретных параметров покажет, в каком конкретно качестве этот нейрон близок к «идеалу».
Далее, в контексте энергетической эффективности нейронов рассмотрены два процесса, на которых основано кодирование и передача информации в мозге. Это нервный импульс, или потенциал действия, благодаря которому информация может быть отправлена «адресату» на определенное расстояние (от микрометров до полутора метров) и синаптическая передача, лежащая в основе собственно передачи сигнала от одного нейрона на другой.
Потенциал действия
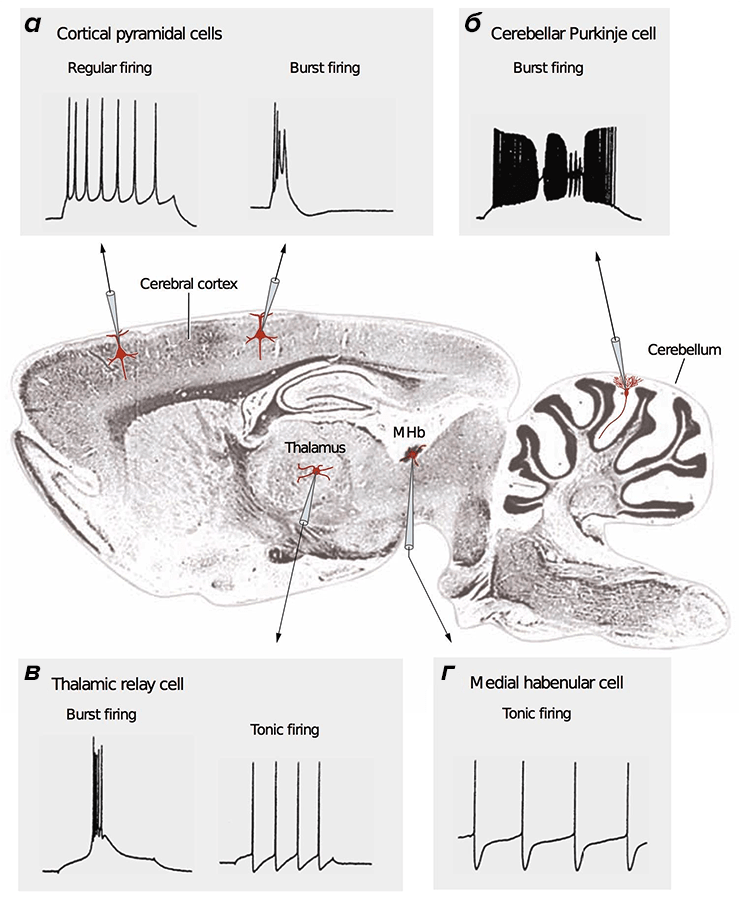
Потенциал действия (ПД) — сигнал, которые отправляют друг другу нейроны. ПД бывают разные: быстрые и медленные, малые и большие [28]. Зачастую они организованы в длинные последовательности (как буквы в слова), либо в короткие высокочастотные «пачки» (рис. 2).
Большое разнообразие сигналов обусловлено огромным количеством комбинаций разных типов ионных каналов, синаптических контактов, а также морфологией нейронов [28], [29]. Поскольку в основе сигнальных процессов нейрона лежат ионные токи, стоит ожидать, что разные ПД требуют различных энергозатрат [20], [27], [30].
Что такое потенциал действия?
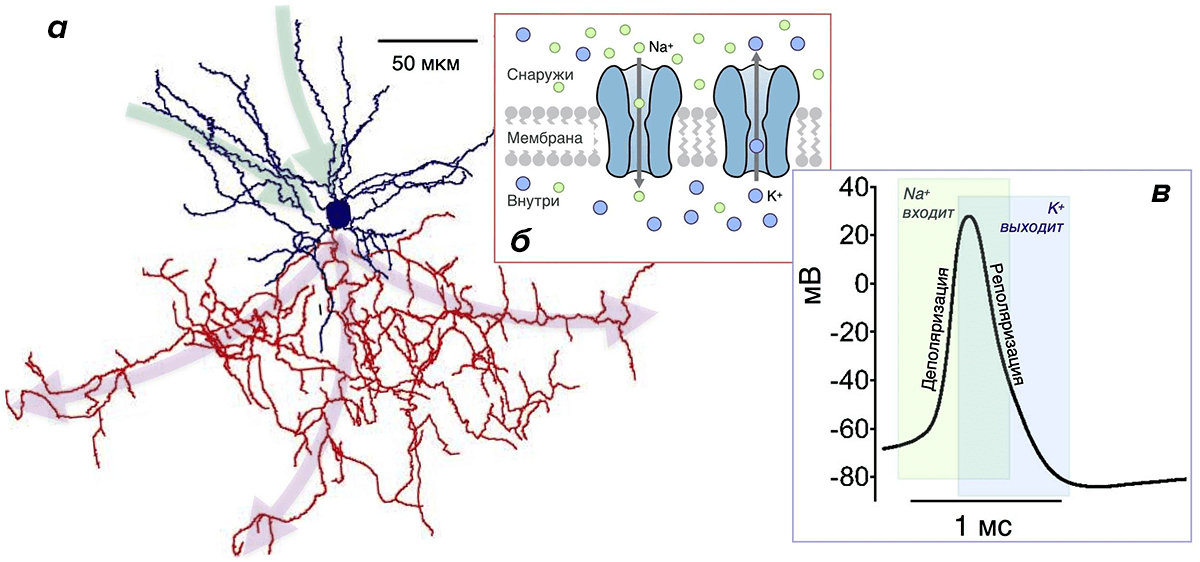
ПД — это относительно сильное по амплитуде скачкообразное изменение мембранного потенциала.
Анализ разных типов нейронов (рис. 4) показал, что нейроны беспозвоночных не очень энергоэффективны, а некоторые нейроны позвоночных почти совершенны [20]. По результатам этого исследования, наиболее энергоэффективными оказались интернейроны гиппокампа, участвующего в формировании памяти и эмоций, а также таламокортикальные релейные нейроны, несущие основной поток сенсорной информации от таламуса к коре больших полушарий.
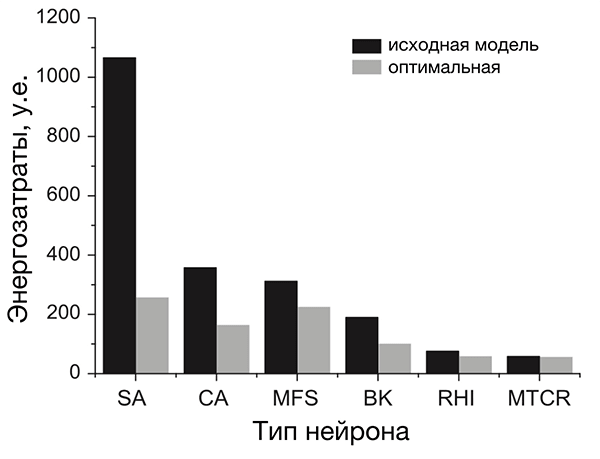
Рисунок 4. Разные нейроны эффективны по-разному. На рисунке представлено сравнение энергозатрат разных типов нейронов. Энергозатраты рассчитаны в моделях как с исходными (реальными) значениями параметров (черные столбцы), так и с оптимальными, при которых с одной стороны нейрон выполняет положенную ему функцию, с другой — затрачивает при этом минимум энергии (серые столбцы). Самыми эффективными из представленных оказались два типа нейронов позвоночных: интернейроны гиппокампа (rat hippocampal interneuron, RHI) и таламокортикальные нейроны (mouse thalamocortical relay cell, MTCR), так как для них энергозатраты в исходной модели наиболее близки к энергозатратам оптимизированной. Напротив, нейроны беспозвоночных менее эффективны. Условные обозначения: SA (squid axon) — гигантский аксон кальмара; CA (crab axon) — аксон краба; MFS (mouse fast spiking cortical interneuron) — быстрый кортикальный интернейрон мыши; BK (honeybee mushroom body Kenyon cell) — грибовидная клетка Кеньона пчелы.
Почему они более эффективны? Потому что у них малó перекрывание Na- и К-токов. Во время генерации ПД всегда есть промежуток времени, когда эти токи присутствуют одновременно (рис. 3в). При этом переноса заряда практически не происходит, и изменение мембранного потенциала минимально. Но «платить» за эти токи в любом случае приходится, несмотря на их «бесполезность» в этот период. Поэтому его продолжительность определяет, сколько энергетических ресурсов растрачивается впустую. Чем он короче, тем более эффективно использование энергии [20], [26], [30], [43]. Чем длиннее — тем менее эффективно. Как раз в двух вышеупомянутых типах нейронов, благодаря быстрым ионным каналам, этот период очень короткий, а ПД — самые эффективные [20].
Кстати, интернейроны гораздо более активны, чем большинство других нейронов мозга. В то же время они крайне важны для слаженной, синхронной работы нейронов, с которыми образуют небольшие локальные сети [9], [16]. Вероятно, высокая энергетическая эффективность ПД интернейронов является некой адаптацией к их высокой активности и роли в координации работы других нейронов [20].
Синапс
Передача сигнала от одного нейрона к другому происходит в специальном контакте между нейронами, в синапсе [12]. Мы рассмотрим только химические синапсы (есть еще электрические), поскольку они весьма распространены в нервной системе и важны для регуляции клеточного метаболизма, доставки питательных веществ [5].
Чаще всего, химический синапс образован между окончанием аксона одного нейрона и дендритом другого. Его работа напоминает. «переброс» эстафетной палочки, роль которой и играет нейромедиатор — химический посредник передачи сигнала [12], [42], [44–48].
На пресинаптическом окончании аксона ПД вызывает выброс нейромедиатора во внеклеточную среду — к принимающему нейрону. Последний только этого и ждет с нетерпением: в мембране дендритов рецепторы — ионные каналы определенного типа — связывают нейромедиатор, открываются и пропускают через себя разные ионы. Это приводит к генерации маленького постсинаптического потенциала (ПСП) на мембране дендрита. Он напоминает ПД, но значительно меньше по амплитуде и происходит за счет открывания других каналов. Множество этих маленьких ПСП, каждый от своего синапса, «сбегаются» по мембране дендритов к телу нейрона (зеленые стрелки на рис. 3а) и достигают начального сегмента аксона, где вызывают открывание Na-каналов и «провоцируют» его на генерацию ПД.
Такие синапсы называются возбуждающими: они способствуют активации нейрона и генерации ПД. Существуют также и тормозящие синапсы. Они, наоборот, способствуют торможению и препятствуют генерации ПД. Часто на одном нейроне есть и те, и другие синапсы. Определенное соотношение между торможением и возбуждением важно для нормальной работы мозга, формирования мозговых ритмов, сопровождающих высшие когнитивные функции [49].
Как это ни странно, выброс нейромедиатора в синапсе может и не произойти вовсе — это процесс вероятностный [18], [19]. Нейроны так экономят энергию: синаптическая передача и так обусловливает около половины всех энергозатрат нейронов [25]. Если бы синапсы всегда срабатывали, вся энергия пошла бы на обеспечение их работы, и не осталось бы ресурсов для других процессов. Более того, именно низкая вероятность (20–40%) выброса нейромедиатора соответствует наибольшей энергетической эффективности синапсов. Отношение количества полезной информации к затрачиваемой энергии в этом случае максимально [18], [19]. Так, выходит, что «неудачи» играют важную роль в работе синапсов и, соответственно, всего мозга. А за передачу сигнала при иногда «не срабатывающих» синапсах можно не беспокоиться, так как между нейронами обычно много синапсов, и хоть один из них да сработает.
Еще одна особенность синаптической передачи состоит в разделении общего потока информации на отдельные компоненты по частоте модуляции приходящего сигнала (грубо говоря, частоте приходящих ПД) [50]. Это происходит благодаря комбинированию разных рецепторов на постсинаптической мембране [38], [50]. Некоторые рецепторы активируются очень быстро: например, AMPA-рецепторы (AMPA происходит от α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid). Если на постсинаптическом нейроне представлены только такие рецепторы, он может четко воспринимать высокочастотный сигнал (такой, как, например, на рис. 2в). Ярчайший пример — нейроны слуховой системы, участвующие в определении местоположения источника звука и точном распознавании коротких звуков типа щелчка, широко представленных в речи [12], [38], [51]. NMDA-рецепторы (NMDA — от N—methyl-D—aspartate) более медлительны. Они позволяют нейронам отбирать сигналы более низкой частоты (рис. 2г), а также воспринимать высокочастотную серию ПД как нечто единое — так называемое интегрирование синаптических сигналов [14]. Есть еще более медленные метаботропные рецепторы, которые при связывании нейромедиатора, передают сигнал на цепочку внутриклеточных «вторичных посредников» для подстройки самых разных клеточных процессов. К примеру, широко распространены рецепторы, ассоциированные с G-белками. В зависимости от типа они, например, регулируют количество каналов в мембране или напрямую модулируют их работу [14].
Различные комбинации быстрых AMPA-, более медленных NMDA- и метаботропных рецепторов позволяют нейронам отбирать и использовать наиболее полезную для них информацию, важную для их функционирования [50]. А «бесполезная» информация отсеивается, она не «воспринимается» нейроном. В таком случае не приходится тратить энергию на обработку ненужной информации. В этом и состоит еще одна сторона оптимизации синаптической передачи между нейронами.
Что еще?
Энергетическая эффективность клеток мозга исследуется также и в отношении их морфологии [35], [52–54]. Исследования показывают, что ветвление дендритов и аксона не хаотично и тоже экономит энергию [52], [54]. Например, аксон ветвится так, чтобы суммарная длина пути, который проходит ПД, была наименьшей. В таком случае энергозатраты на проведение ПД вдоль аксона минимальны.
Снижение энергозатрат нейрона достигается также при определенном соотношении тормозящих и возбуждающих синапсов [55]. Это имеет прямое отношение, например, к ишемии (патологическому состоянию, вызванному нарушением кровотока в сосудах) головного мозга. При этой патологии, вероятнее всего, первыми выходят из строя наиболее метаболически активные нейроны [9], [16]. В коре они представлены ингибиторными интернейронами, образующими тормозящие синапсы на множестве других пирамидальных нейронов [9], [16], [49]. В результате гибели интернейронов, снижается торможение пирамидальных. Как следствие, возрастает общий уровень активности последних (чаще срабатывают активирующие синапсы, чаще генерируются ПД). За этим немедленно следует рост их энергопотребления, что в условиях ишемии может привести к гибели нейронов.
При изучении патологий внимание уделяют и синаптической передаче как наиболее энергозатратному процессу [19]. Например, при болезнях Паркинсона [56], Хантингтона [57], Альцгеймера [58–61] происходит нарушение работы или транспорта к синапсам митохондрий, играющих основную роль в синтезе АТФ [62], [63]. В случае болезни Паркинсона, это может быть связано с нарушением работы и гибелью высоко энергозатратных нейронов черной субстанции, важной для регуляции моторных функций, тонуса мышц. При болезни Хантингтона, мутантный белок хангтингтин нарушает механизмы доставки новых митохондрий к синапсам, что приводит к «энергетическому голоданию» последних, повышенной уязвимости нейронов и избыточной активации. Все это может вызвать дальнейшие нарушения работы нейронов с последующей атрофией полосатого тела и коры головного мозга. При болезни Альцгеймера нарушение работы митохондрий (параллельно со снижением количества синапсов) происходит из-за отложения амилоидных бляшек. Действие последних на митохондрии приводит к окислительному стрессу, а также к апоптозу — клеточной гибели нейронов.
Еще раз обо всем
В конце ХХ века зародился подход к изучению мозга, в котором одновременно рассматривают две важные характеристики: сколько нейрон (или нейронная сеть, или синапс) кодирует и передает полезной информации и сколько энергии при этом тратит [6], [18], [19]. Их соотношение является своего рода критерием энергетической эффективности нейронов, нейронных сетей и синапсов.
Использование этого критерия в вычислительной нейробиологии дало существенный прирост к знаниям относительно роли некоторых явлений, процессов [6], [18–20], [26], [30], [43], [55]. В частности, малая вероятность выброса нейромедиатора в синапсе [18], [19], определенный баланс между торможением и возбуждением нейрона [55], выделение только определенного рода приходящей информации благодаря определенной комбинации рецепторов [50] — все это способствует экономии ценных энергетических ресурсов.
Более того, само по себе определение энергозатрат сигнальных процессов (например, генерация, проведение ПД, синаптическая передача) позволяет выяснить, какой из них пострадает в первую очередь при патологическом нарушении доставки питательных веществ [10], [25], [56]. Так как больше всего энергии требуется для работы синапсов, именно они первыми выйдут из строя при таких патологиях, как ишемия, болезни Альцгеймера и Хантингтона [19], [25]. Схожим образом определение энергозатрат разных типов нейронов помогает выяснить, какой из них погибнет раньше других в случае патологии. Например, при той же ишемии, в первую очередь выйдут из строя интернейроны коры [9], [16]. Эти же нейроны из-за интенсивного метаболизма — наиболее уязвимые клетки и при старении, болезни Альцгеймера и шизофрении [16].
В общем, подход к определению энергетически эффективных механизмов работы мозга является мощным направлением для развития и фундаментальной нейронауки, и ее медицинских аспектов [5], [14], [16], [20], [26], [55], [64].
Благодарности
Искренне благодарен моим родителям Ольге Наталевич и Александру Жукову, сестрам Любе и Алене, моему научному руководителю Алексею Браже и замечательным друзьям по лаборатории Эвелине Никельшпарг и Ольге Слатинской за поддержку и вдохновение, ценные замечания, сделанные при прочтении статьи. Я также очень благодарен редактору статьи Анне Петренко и главреду «Биомолекулы» Антону Чугунову за пометки, предложения и замечания.
Нервная система
%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7/%D0%BF%D0%BE%D1%81%D0%BE%D0%B1%D0%B8%D0%B5%20%D0%90%D0%9D%D0%A1%202016%20(%D0%A7%D0%B0%D1%81%D1%82%D1%8C%20I)%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7..files/image001.gif) | %20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7/%D0%BF%D0%BE%D1%81%D0%BE%D0%B1%D0%B8%D0%B5%20%D0%90%D0%9D%D0%A1%202016%20(%D0%A7%D0%B0%D1%81%D1%82%D1%8C%20I)%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7..files/image002.gif) |
Центральная нервная система
Периферическая нервная система
%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7/%D0%BF%D0%BE%D1%81%D0%BE%D0%B1%D0%B8%D0%B5%20%D0%90%D0%9D%D0%A1%202016%20(%D0%A7%D0%B0%D1%81%D1%82%D1%8C%20I)%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7..files/image003.gif)
%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7/%D0%BF%D0%BE%D1%81%D0%BE%D0%B1%D0%B8%D0%B5%20%D0%90%D0%9D%D0%A1%202016%20(%D0%A7%D0%B0%D1%81%D1%82%D1%8C%20I)%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7..files/image004.gif)
%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7/%D0%BF%D0%BE%D1%81%D0%BE%D0%B1%D0%B8%D0%B5%20%D0%90%D0%9D%D0%A1%202016%20(%D0%A7%D0%B0%D1%81%D1%82%D1%8C%20I)%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7..files/image005.gif)
%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7/%D0%BF%D0%BE%D1%81%D0%BE%D0%B1%D0%B8%D0%B5%20%D0%90%D0%9D%D0%A1%202016%20(%D0%A7%D0%B0%D1%81%D1%82%D1%8C%20I)%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7..files/image006.gif)
%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7/%D0%BF%D0%BE%D1%81%D0%BE%D0%B1%D0%B8%D0%B5%20%D0%90%D0%9D%D0%A1%202016%20(%D0%A7%D0%B0%D1%81%D1%82%D1%8C%20I)%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7..files/image007.gif)
Нейрон имеет отростчатое строение и состоит из тела и отростков (Рис.1). Размеры нейронов колеблются от 6 до 120 мкм. У беспозвоночных, например у моллюсков, размер нейрона может достигать до 300-500 микрон.
%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7/%D0%BF%D0%BE%D1%81%D0%BE%D0%B1%D0%B8%D0%B5%20%D0%90%D0%9D%D0%A1%202016%20(%D0%A7%D0%B0%D1%81%D1%82%D1%8C%20I)%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7..files/image008.jpg)
Рис.1. Внешнее строение нейрона
%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7/%D0%BF%D0%BE%D1%81%D0%BE%D0%B1%D0%B8%D0%B5%20%D0%90%D0%9D%D0%A1%202016%20(%D0%A7%D0%B0%D1%81%D1%82%D1%8C%20I)%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7..files/image010.gif)
Рис.2. Внутреннее строение нейрона
Аксонный холмик – область перикариона, содержащая много микротрубочек и нейрофиламентов, место, где начинается аксон (нейрит) и генерируется потенциал действия.
На оболочке волокна располагаются и так называемые насечки Лантермана, проникающие до самого осевого цилиндра. Эти насечки в толстых волокнах обнаруживаются чаще, чем в тонких. Насечки Лантермана служат для обмена веществ между осевыми цилиндрами и окружающей средой.
Проксимальные области дендритов – продолжение перикариона. Поэтому они содержат рибосомы, компоненты гранулярной и гладкой ЭПС, элементы комплекса Гольджи. Дендриты интегрально образуют до 95% всей рецепторной поверхности (рецептивного поля) нейрона. Они растут и подвергаются ретракции (сокращению и уменьшению в размерах) в ответ на различные воздействия. Дендриты являются наиболее чувствительными индикаторами функционального состояния нервных связей и любых изменений в нервной системе. Дендриты в отличие от аксонов способны генерировать потенциалы действия.
Дендриты и аксоны сохраняют свою форму благодаря микротрубочкам, которые, по-видимому, играют также роль в передвижении синтезированных продуктов из центральной цитоплазмы к очень далеким от нее концам ветвящихся аксонов и дендритов. Аксон и дендриты хотя и имеют в определенной степени сходное строение, но существенно различаются по функциям: аксон проводит импульс от перикариона, а дендриты – к перикариону. Основная часть нейрита, называемая осевым цилиндром, в отличие от дендрита не содержит тигроида. Возбуждение по нейрону проходит от дендрита к перикариону и от него по аксону. В противоположном направлении нервные импульсы не проходят, а тормозятся.
Цитоскелет нейронов состоит из микротрубочек, промежуточных филаментов (нейрофиламенты) и микрофиламентов. микротрубочки – наиболее крупные элементы цитоскелета, их диаметр достигает 24 нм. С ними связывают внутриклеточный, в т.ч. аксонный, транспорт. Микротрубочки обеспечивают структурную жесткость, а также транспортировку материалов, синтезируемых в теле клетки и предназначенных для использовании в окончании аксона (внизу). Микротрубочки в перикарионе и дендритах (в отличие от аксона) не имеют направленной ориентации. Большинство микротрубочек аксона положительным концом направлено к терминали, а отрицательным концом – к перикариону. Характер ориентации микротрубочек имеет важное значение для распределения по отросткам различных органелл. К (+) полюсу перемещаются митохондрии и секреторные пузырьки, а к (-) концу – рибосомы, мультивезикулярные тельца, элементы комплекса Гольджи.
%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7/%D0%BF%D0%BE%D1%81%D0%BE%D0%B1%D0%B8%D0%B5%20%D0%90%D0%9D%D0%A1%202016%20(%D0%A7%D0%B0%D1%81%D1%82%D1%8C%20I)%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7..files/image011.jpg)
Нейроны между собой отличаются по размерам, форме перикариона, числом отростков, их синаптическими связями, характером ветвления дендритов, электрофизиологическими характеристиками, химией нейромедиаторов, позицией в функциональных сетях и т.д. в связи с этим классификации нейронов многочисленны.
Морфологическая характеристика нейрона определяется количеством отходящих от него отростков. По этому признаку различают нейроны: мультиполярные с большим количеством отростков; биполярные – с двумя отростками; униполярные – с одним отростком. Униполярные и биполярные нейроны округлые или овальные, а мультиполярные имеют неправильную многоугольную форму (Рис.3).
%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7/%D0%BF%D0%BE%D1%81%D0%BE%D0%B1%D0%B8%D0%B5%20%D0%90%D0%9D%D0%A1%202016%20(%D0%A7%D0%B0%D1%81%D1%82%D1%8C%20I)%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7..files/image013.jpg)
Рис. 3. Морфологическая классификация нейронов
%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7/%D0%BF%D0%BE%D1%81%D0%BE%D0%B1%D0%B8%D0%B5%20%D0%90%D0%9D%D0%A1%202016%20(%D0%A7%D0%B0%D1%81%D1%82%D1%8C%20I)%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7..files/image015.jpg)
%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7/%D0%BF%D0%BE%D1%81%D0%BE%D0%B1%D0%B8%D0%B5%20%D0%90%D0%9D%D0%A1%202016%20(%D0%A7%D0%B0%D1%81%D1%82%D1%8C%20I)%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7..files/image017.jpg)
б.Мультиполярные нервные клетки. Гистологический препарат передних рогов спинного мозга. Окраска нигрознином Ув.:об.8×ок.15. 1-мультиполярная нервная клетка (нейроцит) спинного мозга;2-ядро нервной клетки;3 – дендриты; 4 – аксон (нейрит).
Мультиполярные нейроны располагают числом отростков больше, чем 2. Среди этих отростков имеется 1 аксон, а остальные – дендриты. Насчитывают до 60 различных вариантов строения мультиполярных нейронов, однако все они представляют разновидности веретенообразных, звездчатых, корзинчатых и пирамидных клеток. Примером мультиполярного нейрона могут служить мотонейроны передних рогов спинного мозга (Рис.4б).
По соотношению размера ядра и окружающей его цитоплазмы различают кариохромные и соматохромные нейроны. Кариохромные нейроны характеризуются тем, что обладают крупным ядром, окруженным узким ободком цитоплазмы. У соматохромного нейрона слой цитоплазмы, окружающий ядро, хорошо выражен.
Вставочные (замыкательные, кондукторные, промежуточные), служащие для переработки и переключения импульсов. Один или несколько вставочных нейронов могут находиться между афферентным и эфферентным нейронами. Вставочные нейроны наиболее многочисленны и расположены во всех отделах спинного и головного мозга. ЦНС на 90% состоит из вставочных нейронов.
Существует также классификация по признаку положения в сети нейронов относительно места действия: первичные, вторичные, третичные и т.д.
Нейроны различаются между собой и размерами отростков. Нейроны с длинными аксонами – это клетки Гольджи 1-го типа, а нейроны с короткими аксонами – клетки Гольджи 2-го типа. В рамках данной классификации короткими считаются такие аксоны, ветви которых находятся в непосредственной близости от тела клетки. Клетки Гольджи 1-го типа (эфферентные) – нейроны с длинным аксоном, продолжающимся в белом веществе мозга.
Кроме того, в зависимости от локализации различают следующие виды нервных окончаний – рецепторов: экстерорецепторы, интерорецепторы и проприорецепторы. Первые воспринимают раздражения, идущие из внешней среды при контакте или на расстоянии. Интерорецепторы воспринимают раздражения из внутренних органов. Среди них различают терморецепторы, механорецепторы, хеморецепторы, барорецепторы, ноцирецепторы (болевые).
б) адренэргические. Нейромедиатор – норадреналин. К ним относятся постганглионарные нейроны симпатического отдела вегетативной нервной системы, иннервирующие сердце, ГМК сосудов и внутренних органов.
%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7/%D0%BF%D0%BE%D1%81%D0%BE%D0%B1%D0%B8%D0%B5%20%D0%90%D0%9D%D0%A1%202016%20(%D0%A7%D0%B0%D1%81%D1%82%D1%8C%20I)%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7..files/image019.jpg)
А – веретеновидный нейрон; Б – пирамидальный нейрон; В – клетка Пуркинье; Г – звездчатый нейрон.
Рис.5. Классификация нейронов по форме тела и ветвлению отростков
Пирамидные нервные клетки встречаются во всех слоях коры больших полушарий. Они сильно варьируют по своим размерам. Крупные нейроны (гигантопирамидные) располагаются в IV слое зрительной коры, а также в III и I V слоях корковых зон анализаторов. Наиболее крупные нейроны, известные как клетки Беца В.А., обнаруживаются в области коркового конца двигательного анализатора. В местах деления III на три подслоя гигантопирамидные нейроны залегают в третьем подслое.
Бисенсорные нейроны. Чаще располагаются во вторичных зонах коры какого-либо анализатора и могут реагировать на сигналы как своей, так и другой сенсорности. Например, нейроны вторичной зоны зрительной области коры большого мозга реагируют на зрительные и слуховые раздражения.
Полисенсорные нейроны. Это чаще всего нейроны ассоциативных зон мозга; они способны реагировать на раздражение слуховой, зрительной, кожной и других рецептивных систем.
Специфические образования нервной клетки. К специфическим образованиям относятся тигроидное вещество и нейрофибриллы. Тигроидное вещество (тигроид, вещество Ниссля) находится в перикарионе и дендритах, он отсутствует в аксоне. Под световым микроскопом тигроид выявляется как скопление базофильного вещества в виде глыбок или зерен. Крупные глыбки придают цитоплазме пятнистый вид шкуры тигра. С помощью электронного микроскопа установлено, что тигроид представляет мощно развитый гранулярный ЭПР. Ретикулум состоит из системы мембран с большим количеством рибосом. Высокое содержание РНК обуславливает базофилию тигроида. В нем содержится и белок.
Тигроид – обязательный компонент нервной клетки, легко меняющийся в зависимости от функционального состояния. Тигролиз – распыление тигроидного вещества, отражает глубокие дистрофические изменения при нарушении целостности нейронов. При сильном возбуждении нейрона тигроид может исчезнуть вообще. Уменьшение тигроида и изменение его положения в нейронах наблюдается также в результате патологических процессов: воспаления, дегенерации, интоксикации. Все это дает основание рассматривать количество тигроида, форму его глыбок, характер их расположения как показатели физиологического состояния нейрона.
В цитоплазме нейронов обнаруживаются нейрофибриллы – нитчатые структуры. В теле нейрона и дендритах они образуют густую сеть. В аксоне они вытягиваются по длине. Открытие нейрофибрилл привело к возникновению нейрофибриллярной теории проведения нервного возбуждения. Сторонники этой теории считали, что нейрофибриллы являются беспрерывным проводящим элементом нервной системы, с чем связана ее главная функция. В дальнейшем было установлено, что нейрофибриллы не принимают участие в процессе проведения нервного и возбуждения и прерываются в области контакта нервных клеток.
По современным представлениям, в соответствии с нейронной теорией в проведении нервного возбуждения основная роль принадлежит плазмалемме нейрона. Вопрос о значении фибрилл остается неясным. По слипанию нейрофибрилл определяют патологическое состояние нервной клетки. Показано, что при старческом слабоумии наблюдается слипание и огрубление нейрофибриллярной сети.
Липиды нейронов служат энергетическим и пластическим материалом. Присутствие в миелиновой оболочке липидов обусловливает их высокое электрическое сопротивление. Обмен липидов в нервной клетке происходит медленно; возбуждение нейрона приводит к уменьшению количества липидов. Обычно после длительной умственной работы, при утомлении количество фосфолипидов в клетке уменьшается.
Углеводы нейронов являются основным источником энергии для них. Глюкоза, поступая в нервную клетку, превращается в гликоген, который при необходимости под влиянием ферментов самой клетки превращается вновь в глюкозу. Вследствие того, что запасы гликогена при работе нейрона не обеспечивают полностью его энергетические траты, источником энергии для нервной клетки служит и глюкоза крови. Расщепление глюкозы идет преимущественно аэробным путем, чем объясняется высокая чувствительность нервных клеток к недостатку кислорода. Увеличение в крови адреналина, активная деятельность организма приводят к увеличению потребления углеводов.
Обмен энергии в нейроне в состоянии покоя и возбуждения различен. После возбуждения количество нуклеиновых кислот в цитоплазме нейронов иногда уменьшается в 5 раз. Собственные энергетические процессы нейрона (его сомы) тесно связаны с трофическими влияниями нейронов, что сказывается, прежде всего, на аксонах и дендритах. В то же время нервные окончания аксонов оказывают трофические влияния на мышцу или клетки других органов. Так, нарушение иннервации мышцы приводит к ее атрофии, усилению распада белков, гибели мышечных волокон.
Тема 3. Нейросекреторные клетки. Регенерация нейронов. Нейроглия.
Нейросекреторные нервные клетки. В определенных отделах мозга беспозвоночных и позвоночных животных имеются нейроны, содержащие гранулы секрета. Такие секретирующие нейроны называются нейросекреторными. Они имеют физиологические признаки нейрона, но обладают выраженными признаками железистых клеток. Нейросекрет синтезируются в связи с тигроидной субстанцией гранулярной ЭПС, оформляется в виде секрета в системе аппарата Гольджи. Секрет продвигается по аксону и выделяется из клеток в области их концевых разветвлений. В отличие от обычных нейронов секрет высвобождается не в области синапса, а в кровь или ликвор (мозговую жидкость). Аксоны нейросекреторных клеток направляется в нейрогипофиз и промежуточную долю аденогипофиза, образуя с ними единую систему. Выделяемый нейросекреторными клетками продукт рассматривают как гормон, регулирующий деятельность некоторых желез внутренней секреции и гонад, где нервная регуляция оказывается редуцированной.
Регенерация нейронов. Природа закладывает в развивающийся мозг очень высокий запас прочности: при эмбриогенезе образуется большой избыток нейронов. Почти 70% из них гибнут еще до рождения ребенка. Человеческий мозг продолжает терять нейроны и после рождения, на протяжении всей жизни. Такая гибель клеток генетически запрограммирована. Как же люди умудряются сохранить интеллект до весьма преклонных лет, если нервные клетки погибают и не обновляются?
Смысл пластичности в том, что функции погибших нервных клеток берут на себя их оставшиеся в живых нервные клетки, которые увеличиваются в размерах и формируют новые связи, компенсируя утраченные функции. Высокую, но не беспредельную эффективность подобной компенсации можно проиллюстрировать на примере болезни Паркинсона, при которой происходит постепенное отмирание нейронов. Оказывается, пока в головном мозге не погибнет около 90% нейронов, клинические симптомы заболевания (дрожание конечностей, ограничение подвижности, неустойчивая походка, слабоумие) не проявляются, то есть человек выглядит практически здоровым. Значит, одна живая нервная клетка может заменить девять погибших.
Первое сообщение о нейрогенезе появилось в 1962 году в статье «Формируются ли новые нейроны в мозге взрослых млекопитающих?». Ее автор, профессор Ж. Олтман. Он с помощью электрического тока разрушал латеральное коленчатое тело крысы и вводил туда радиоактивное вещество, проникающее во вновь возникающие клетки. Через несколько месяцев ученый обнаружил новые радиоактивные нейроны в таламусе и коре головного мозга.
В дальнейшем аналогичное явление было установлено и другими исследователями в головном мозге птиц. В конце 1980-х годов нейрогенез был также обнаружен у взрослых амфибий в лаборатории ленинградского ученого профессора А. Л. Поленова.
%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7/%D0%BF%D0%BE%D1%81%D0%BE%D0%B1%D0%B8%D0%B5%20%D0%90%D0%9D%D0%A1%202016%20(%D0%A7%D0%B0%D1%81%D1%82%D1%8C%20I)%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7..files/image021.jpg)
Рис.6. Основные пути дифференцировки клеток ганглионарной пластинки и нервной трубки
Развитие нейробиологии в начале 1990-х годов привело к обнаружению «новорожденных» нейронов в головном мозге взрослых крыс и мышей. Их находили большей частью в эволюционно древних отделах головного мозга: обонятельных луковицах и коре гиппокампа, которые отвечают главным образом за эмоциональное поведение, реакцию на стресс и регуляцию половых функций млекопитающих.
Стволовые клетки можно извлечь из мозга и пересадить в другой участок нервной системы, где они превратятся в нейроны. Профессор Гейдж с коллегами провел несколько подобных экспериментов, наиболее впечатляющим среди которых был следующий. Участок мозговой ткани, содержащий стволовые клетки, пересадили в разрушенную сетчатку глаза крысы. Пересаженные стволовые клетки мозга превратились в нейроны сетчатки, их отростки достигли зрительного нерва, и крыса прозрела!
Исследования показали, что новые нейроны во взрослом мозге могут образовываться не только из нейрональных стволовых клеток, но и из стволовых клеток крови. Оказалось, что стволовые клетки действительно проникают в мозг, но они не превращаются в нейроны, а сливаются с ними, образую двуядерные клетки. Затем «старое» ядро нейрона разрушается, а его замещает «новое» ядро стволовой клетки крови. Согласно одной из гипотез, стволовые клетки несут новый генетический материал, который, попадая в «старую» клетки мозжечка, продлевает его жизнь.
Наибольшая сложность в работе с эмбриональными клетками – заставить их трансформироваться в нейроны. Новые технологии позволяют сделать это.
Трансплантация стволовых клеток, несомненно, будет одним из главных подходов в терапии таких нейродегенеративных заболеваний, как болезни Альцгеймера и Паркинсона.
Нейроглия. Термин « нейроглия» ввел в обиход немецкий патологоанатом Рудольф Вирхов для описания связывающих элементов между нейронами. Эти клетки составляют половину объема мозга. Нейроны — это высокоспециализированные клетки, существующие и функционирующие в строго определенной среде. Такую среду им обеспечивает нейроглия. Нейроглия – вспомогательная и очень важная составная часть нервной ткани, связанная с нейронами. По мере специализации нейрона как индивидуальной клетки в процессе эволюции возникла организация более высокого порядка – межклеточное «сообщество» нейрона и нейроглии. Нейроглия не принимает непосредственного участия генерации и проведении нервных импульсов и, тем не менее, нормальное функционирование нейрона невозможно в отсутствии или при повреждении глии. Нейроглия выполняет следующие функции: опорную, трофическую, разграничительную, поддержание постоянства среды вокруг нейронов, защитную, секреторную. Клетки нейроглии не образуют синапсов. Различают глию центральной и периферической нервной системы.
Клетки глии центральной нервной системы делятся на макроглию и микроглию.
Макроглия развивается из глиобластов нервной трубки и включает: эпендиму, астроглию и олигодендроглию. Эпендимоциты выстилают желудочки головного мозга и центральный канал спинного мозга. Эти клетки цилиндрической формы. Они образуют слой типа эпителия, носящий название эпендимы. Между соседними клетками эпендимы имеются щелевидные соединения и пояски сцепления, но плотные соединения отсутствуют, так что цереброспинальная жидкость может проникать между эпендимоцитами в нервную ткань. Большинство эпендимоцитов имеют подвижные реснички, вызывающие ток цереброспинальной жидкости. Базальная поверхность большинства эпендимоцитов ровная, но некоторые клетки имеют длинный отросток, идущий глубоко в нервную ткань. Такие клетки называются таницитами. Они многочисленны в дне III желудочка. Считается, что эти клетки передают информацию о составе цереброспинальной жидкости на первичную капиллярную сеть воротной системы гипофиза. Эпендимный эпителий сосудистых сплетений желудочков продуцирует цереброспинальную жидкость (ликвор).
Протоплазматические астроциты характеризуются короткими сильно ветвящимися отростками и светлым сферическим ядром. Отростки астроцитов тянутся к базальным мембранам капилляров, к телам и дендритам нейронов, окружая синапсы и отделяя (изолируя) их друг от друга, а также к мягкой мозговой оболочке, образуя пиоглиальную мембрану, граничащую с субарахноидальным пространством. Подходя к капиллярам, их отростки образуют расширенные «ножки», полностью окружающие сосуд. Астроциты накапливают и передают вещества от капилляров к нейронам, захватывают избыток экстрацеллюлярного калия и других веществ, таких как нейромедиаторы, из экстрацеллюлярного пространства после интенсивной нейрональной активности.
Олигодендроглию образуют олигодендроциты. Олигодендроциты – имеют более мелкие по сравнению с астроцитами и более интенсивно окрашивающиеся ядра. Их отростки немногочисленны. Олигодендроглиоциты присутствуют как в сером, так и в белом веществе. В сером веществе они локализуются вблизи перикарионов. В белом веществе их отростки образуют миелиновый слой в миелиновых нервных волокнах, причем, в противоположность аналогичным клеткам периферической нервной системы – нейролеммоцитам, один олигодендроглиоцит может участвовать в миелинизации сразу нескольких аксонов.
Микроглия образуют микроглиоциты, которые представляют собой фагоцитирующие клетки, относящиеся к системе мононуклеарных фагоцитов и происходящие из стволовой кроветворной клетки (возможно, из премоноцитов красного костного мозга). Функция микроглии — защита от инфекции и повреждения, и удаление продуктов разрушения нервной ткани. Клетки микроглии характеризуются небольшими размерами, телами продолговатой формы. Их короткие отростки имеют на своей поверхности вторичные и третичные ответвления, что придает клеткам «колючий» вид. Описанная морфология характерна для типичной (ветвистой, или покоящейся) микроглии полностью сформированной центральной нервной системы. Она обладает слабой фагоцитарной активностью. Ветвистая микроглия встречается как в сером, так и в белом веществе центральной нервной системы. В белом веществе мозга на долю клеток микроглии приходится 5% от всех видов клеток, а в сером веществе их доля составляет 18%.
В развивающемся мозгу млекопитающих обнаруживается временная форма микроглии — амебоидная микроглия. Клетки амебоидной микроглии формируют выросты – филоподии и складки плазмолеммы. В их цитоплазме присутствуют многочисленные фаголизосомы и пластинчатые тельца. Амебоидные микроглиальные тельца отличаются высокой активностью лизосомальных ферментов. Активно фагоцитирующая амебоидная микроглия необходима в раннем постнатальном периоде, когда гематоэнцефалический барьер еще не вполне развит и вещества из крови легко попадают в центральную нервную систему. Считают также, что она способствует удалению обломков клеток, появляющихся в результате запрограммированной гибели избыточных нейронов и их отростков в процессе дифференцировки нервной системы. Полагают, что, созревая, амебоидные микроглиальные клетки превращаются в ветвистую микроглию.
Реактивная микроглия появляется после травмы в любой области мозга. Она не имеет ветвящихся отростков, как покоящаяся микроглия, не имеет псевдоподий и филоподий, как амебоидная микроглия. В цитоплазме клеток реактивной микроглии присутствуют плотные тельца, липидные включения, лизосомы. Есть данные о том, что реактивная микроглия формируется вследствие активации покоящейся микроглии при травмах центральной нервной системы.
Рассмотренные выше глиальные элементы относятся к центральной нервной системе.
Глия периферической нервной системы в отличие от макроглии центральной нервной системы происходит из нервного гребня. К периферической нейроглии относятся: нейролеммоциты (или шванновские клетки) и глиоциты ганглиев (или мантийные глиоциты). Нейролеммоциты и шванновские клетки формируют оболочки отростков нервных клеток в нервных волокнах периферической нервной системы. Мантийные глиоциты ганглиев окружают тела нейронов в нервных узлах и участвуют в обмене веществ этих нейронов.
В отличие от нейронов нейроглия содержит малодифференцированные клетки способные к регенерации, размножению и развитию в течении всей жизни.
Тема 4. Нервные узлы. Нервные волокна. Нервные стволы (нервы)
Спинномозговой узел (спинальный ганглий ). Спинномозговой узел имеет веретеновидную форму, окружен капсулой из плотной соединительной ткани. От капсулы в паренхиму узла проникают тонкие прослойки соединительной ткани, в которой расположены кровеносные сосуды. Нейроны спинномозгового узла характеризуются крупным сферическим телом и светлым ядром с хорошо заметным ядрышком. Клетки располагаются группами, преимущественно по периферии органа. Центр спинномозгового узла состоит главным образом из отростков нейронов и тонких прослоек эндоневрия, несущих сосуды. Дендриты нервных клеток идут в составе чувствительной части смешанных спинномозговых нервов на периферию и заканчиваются там рецепторами. Аксоны в совокупности образуют задние корешки, несущие нервные импульсы в спинной мозг или продолговатый мозг.
Дендриты и аксоны клеток в узле и за его пределами покрыты миелиновыми оболочками из нейролеммоцитов. Тело каждой нервной клетки в спинномозговом узле окружено слоем уплощенных клеток олигодендроглии, которые здесь называются мантийными глиоцитами, или глиоцитами ганглия, или же клетками-сателлитами. Они расположены вокруг тела нейрона и имеют мелкие округлые ядра. Снаружи глиальная оболочка нейрона покрыта тонковолокнистой соединительнотканной оболочкой. Клетки этой оболочки отличаются овальной формой ядер. Нейроны спинномозговых узлов содержат такие нейромедиаторы, как ацетилхолин, глутаминовая кислота.
По функциональному признаку и локализации вегетативные нервные узлы разделяют на симпатические и парасимпатические. Большинство внутренних органов имеет двойную вегетативную иннервацию, т.е. получает постганглионарные волокна от клеток, расположенных как в симпатических, так и в парасимпатических узлах. Реакции, опосредуемые их нейронами, часто имеют противоположную направленность (так, например, симпатическая стимуляция усиливает сердечную деятельность, а парасимпатическая ее тормозит).
Тела нейронов образуют серое вещество головного и спинного мозга, а также нервные ганглии беспозвоночных и позвоночных животных. Связь ЦНС и ганглиев с органами осуществляется при помощи проводящих элементов – нервов, основу которых составляют нервные волокна. Нервы, или нервные стволы, связывают нервные центры головного и спинного мозга с рецепторами и рабочими органами, или же с нервными узлами. Отростки нервных клеток, окруженные плазмалеммой олигодендроцитов или шванновских клеток, называются нервными волокнами (рис.7). Отросток нервной клетки в составе нервного волокна называются осевым цилиндром, а глиальные клетки, формирующие оболочку волокна, называются леммоцитами, или шванновскими клетками.
Нервные волокна образуют в головном и спинном мозге проводящие пути, а на периферии – нервы. В пределах ЦНС нервные волокна входят в состав белого вещества мозга. По нервным волокнам осуществляется проведение нервных импульсов.
Безмякотные (немиелиновые) нервные волокна входят в состав периферических нервов, идущих к внутренним органам, но многие сенсорные волокна также являются безмякотными. Они имеют несколько осевых цилиндров (3-5, иногда до 12), окруженных шванновскими клетками.
В электронных микрофотографиях видно, что каждый осевой цилиндр погружен в леммоцит, ее клеточная мембрана смыкается и образует мезаксон – сдвоенные мембраны шванновской клетки. Каждая шванновская клетка подобным образом окружает несколько осевых цилиндров, погруженных в леммоцит, может быть в разное количество мезаксонов в нервном волокне. Миелин отсутствует. Шванновские клетки на всем протяжении окутывают безмякотное волокно, препятствуя его соприкосновению с окружающей средой.
%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7/%D0%BF%D0%BE%D1%81%D0%BE%D0%B1%D0%B8%D0%B5%20%D0%90%D0%9D%D0%A1%202016%20(%D0%A7%D0%B0%D1%81%D1%82%D1%8C%20I)%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7..files/image023.gif)
Рис.7. Строение нерва (А) и нервного волокна (Б).
%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7/%D0%BF%D0%BE%D1%81%D0%BE%D0%B1%D0%B8%D0%B5%20%D0%90%D0%9D%D0%A1%202016%20(%D0%A7%D0%B0%D1%81%D1%82%D1%8C%20I)%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7..files/image025.jpg)
Рис.8. Поперечное строение нерва (а), нервного волокна (б).
Поскольку отростки нервных клеток покрыты плазмалеммой шванновских клеток только один раз, то нервный импульс при прохождении рассеивается. Он проходит по безмякотным нервным волокнам в 10 раз медленнее, по сравнению с мякотными. По безмякотным волокнам импульс проходит со скоростью меньше 1м/с.
Мякотные нервные волокна составляют белое вещество головного и спинного мозга и входят в периферические нервы. Мякотное нервное волокно состоит из одного осевого цилиндра, вокруг которого шванновские клетки образуют миелиновую оболочку. Нервное волокно, состоящее из одного осевого цилиндра и расположенных вокруг него шванновских клеток, называют мякотным, или миелиновым.
Характерная особенность шванновских клеток – наличие в них липоидного вещества миелина, который образует вокруг осевого цилиндра мякотную (миелиновую) оболочку. Каждая шванновская клетка миелинизирует небольшой сегмент только одного аксона. Мякотная, или миелиновая, оболочка примыкает к осевому цилиндру и окружает его чехлом. Она выполняет роль изолятора. Этим объясняется большая скорость проведения нервных импульсов мякотными нервными волокнами, т. к. скорость проведения возбуждения зависит от диаметра и степени миелинизации нервного волокна.
Миелин регулярно прерывается через определенные промежутки. Фактически эти участки, лишенные миелина, являются границами между двумя соседними клетками, где они соединяются при помощи коротких отростков и называются узлами нервного волокна (перехват Ранвье).
Участок между узлами называется межузловым сегментом. в периферических нервах каждый миелиновый сегмент пересекается с воронкообразными щелями, идущими в косом направлении от наружной поверхности оболочки к внутренней. Они называются « насечками миелина» (Шмидтлантермановскими насечками). Шмидтлантермановские насечки – это участки расслоения миелина, образовавшиеся при миелинизации. Функция насечек неясна. В зависимости от длины миелинового сегмента количество насечек миелина бывает различным. Они отсутствуют в пределах ЦНС.
Осевой цилиндр содержит митохондрии, элементы гладкой ЭПС, элементы цитоскелета – микротрубочки, нейрофиламенты и микрофиламенты. Скорость проведения нервного импульса зависит от диаметра аксона, а сам диаметр определяется количеством содержащихся в нем нейрофиламентов. В нормальных и патологических условиях количество нейрофиламентов и диаметр аксона тесно коррелируют.
Аксонный транспорт обеспечивает кинезии микротрубочек. Различают быстрый (100-1000 мм/сутки) и медленный (1-10 мм/сутки) аксонный транспорт, а также антероградный (транспорт от перикариона) и ретроградный (к перикариону). Основной материал антероградного транспорта – белки, синтезированные в перикарионе (например, белки ионных каналов, ферменты синтеза нейромедиаторов).
Внешняя плазмалемма шванновских клеток окружена базальной мембраной. Наличие в мякотном нервном волокне только одного отростка нервной клетки, изолированного миелиновой оболочкой, а также узловых перехватов, обеспечивает быстрое (до 120 м/с) и точное проведение нервных импульсов.
Выше изложено особенности строения мякотного периферического нервного волокна. Мякотные нервные волокна ЦНС построены сходным образом. Однако оболочка их образована не леммоцитами, а олигодендроцитами. Насечки и перехваты в них отсутствуют, нет и базальных мембран.
Периневриальный барьер необходим для поддержания гомеостаза в эндоневрии. Барьер контролирует транспорт молекул через Периневрий к нервным волокнам, предотвращает доступ в эндоневрий инфекционных агентов.
Периферический нерв содержит разветвленную сеть кровеносных сосудов. В эпиневрии и в наружной части периневрия содержатся артериолы и венулы, а также лимфатические сосуды. В эндоневрии проходят кровеносные капилляры. Периферический нерв иннервирован – имеет специальные нервные волокна.
Тема 5. Синапсы. Медиаторы. Нервные сети.
Соединение нервов между собой (синапсы ). Нейроны, как отдельные единицы нервной системы, функционируют не изолированно. Они соединены между собой и образуют единую сеть, которая передает возбуждение от рецепторов в ЦНС и от нее в различные органы (рис.9,10).
%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7/%D0%BF%D0%BE%D1%81%D0%BE%D0%B1%D0%B8%D0%B5%20%D0%90%D0%9D%D0%A1%202016%20(%D0%A7%D0%B0%D1%81%D1%82%D1%8C%20I)%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7..files/image027.gif)
Рис.9. Синаптические входы нейрона. Синаптические бляшки окончаний пресинаптичесиих аксонов образуют соединения на дендритах и теле (соме)- постсинаптического нейрона.
%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7/%D0%BF%D0%BE%D1%81%D0%BE%D0%B1%D0%B8%D0%B5%20%D0%90%D0%9D%D0%A1%202016%20(%D0%A7%D0%B0%D1%81%D1%82%D1%8C%20I)%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7..files/image029.gif)
Рис.10. Схема выброски медиатора и процессов, происходящих в гипотетическом центральном синапсе.
Конечные участки аксонов и дендритов в области синапса не имеют мякотной оболочки и расширены в пресинаптический мешочек. Мешочек характерен для синаптических пузырьков, имеющих диаметр 40-59 нм. В них содержится медиатор. В зависимости от типа выделяемого медиатора различают синапсы:
а) холинэргические – выделяют ацетилхолин;
б) адренэргические – выделяют норадреналин, дофамин (катехоламины);
в) серотонинэргические – выделяют серотонин;
г) пептидэргические – выделяют пептиды (эндорфины, энкефалины) и аминокислоты (глицин, глутамат, ГАМК). В таких синапсах передача нервного импульса осуществляется при помощи химического вещества – медиатора. Такие синапсы называются синапсами с химической передачей. При изменении мембранного потенциала в терминалях нейромедиаторы выходят в синаптическую щель через поры диаметром 4-5 нм, имеющиеся в пресинаптической мембране (экзоцитоз) и связываются со своими рецепторами в постсинаптической мембране, вызывая изменение мембранного потенциала постсинаптического нейрона. Основными медиаторами являются:
1. Ацетилхолин – один из первых выявленных медиатора. Он известен как «вещество блуждающего нерва» из-за своего воздействия на сердечную деятельность.
2. ГАМК – γ-аминомасляная кислота. Представляет собой наиболее распространенный медиатор ЦНС.
3. Аминокислота глицин, оказывающая тормозное действие на мотонейроны.
4. Кислая аминокислота глутамат, является самым распространенным возбуждающим медиатором ЦНС.
5. Адреналин, норадреналин и дофамин – представляют собой семейство медиаторов, передающих возбуждение или торможение в ЦНС, так и в периферической нервной системе. избыточное выделение дофамина – один из признаков развития шизофрении.
Синаптическая передача – сложный каскад событий. Она возможна при реализации ряда последовательных процессов: синтез нейромедиатора, его накопление и хранение в синаптических пузырьках вблизи пресинаптической мембраны, высвобождение нейромедиатора из нервной терминали, кратковременное взаимодействие нейромедиатора с рецептором, встроенным в постсинаптическую мембрану, разрушение нейромедиатора или захват его нервной терминалью. Многие неврологические и психические заболевания сопровождаются нарушениями синаптической передачи.
Медиаторы связываются со специфическими рецепторами постсинаптической мембраны. Вокруг рецептора формируется область высокой концентрации вещества того или иного медиатора. Соответственно повышается или понижается вероятность открывания ионного канала, т.к. изменяется его проводимость. В синапсах возбуждение проводится только в одном направлении, но гораздо медленней, чем по нервному волокну. Однако передача информации осуществляется исключительно точно.
В некоторых синапсах синаптическая щель отсутствует и его структурной основой является плотный контакт. В таком синапсе возбуждение может передаваться без участия медиатора, т.к. мембраны клеток соприкасаются. Эти синапсы называются синапсами с электрической передачей. В синапсах такого строения пресинаптическая мембрана также имеет поры, но они в 5 раз меньше, чем в синапсах с химической передачей возбуждения. Поры электрических синапсов являются межклеточными диффузионными каналами, соединяющими соприкасающиеся клетки.
По структуре и локализации синапсы подразделяются на 3 группы: межнейронные, рецепторно – нейрональные и нейроэффкторные. Межнейронные синапсы подразделяются на аксодендритические, аксосоматические и аксо-аксональные. Межнейронные синапсы являются синапсами между двумя нейронами. Если аксон одного нейрона контактирует с дендритом другого постсинаптического нейрона, то такие синапсы называются аксодендритическими. Аксодендрическая связь представлена синапсами двух типов. Один тип – это синапсы с широкой синаптической щелью и сами мембраны более утолщены. Такие синапсы характерны для возбуждающих нейронов. Другие синапсы принадлежат тормозным нейронам. Если аксон одного нейрона контактирует с перикарионом другого постсинаптического нейрона, то такой синапс называется аксосоматическим. Если же аксон одного нейрона контактирует с аксоном другого постсинаптического нейрона, то такой синапс называется аксо-аксональным.
Межнейронные синапсы очень многочисленны. На поверхности перикариона и отростков одного пирамидного нейрона в коре больших полушарий головного мозга имеется около 10 4 синапсов.
Нейроэффкторные (аксоэффекторные) синапсы являются контактами между аксоном двигательных (эффекторных) нейронов и клетками, не принадлежащими к нервной системе.
У человека и млекопитающих хорошо изучены двигательные и секреторные нейроэффекторные синапсы, или эффекторные нервные окончания. Первые представляют собой синаптические соединения между аксоном двигательного нейрона и поперечнополосатыми мышечными волокнами, поперечнополосатыми и гладкомышечными клетками, а вторые – между аксонами двигательного нейрона с секреторными клетками. Существуют многочисленные синапсы между аксоном эфферентного нейрона и другими клетками – жировыми, ресничными и др.
Нервные сети. Для того чтобы мозг нормально функционировал, потоки нервных сигналов должны находить надлежащие пути среди клеток различных функциональных систем и межрегиональных объединений. Однако до сих пор остается загадкой, каким образом аксоны и дендриты той или иной нервной клетки растут именно в том направлении, чтобы создавались специфические связи, необходимые для ее функционирования.
Высокая специфичность структуры мозга имеет важное значение. Общий диапазон связей для большинства нервных клеток, по-видимому, предопределен заранее, причем эта предопределенность касается тех клеточных свойств, которые ученые считают генетически контролируемыми. Набор генов, предназначенных для проявления в развивающейся нервной клетке, каким-то еще до конца не установленным образом определяет как будущий тип каждой нервной клетки, так и принадлежность ее к той или иной сети. Концепция генетической детерминированности приложима и ко всем остальным особенностям данного нейрона, например к используемому им медиатору, к размерам и форме клетки. Как внутриклеточные процессы, так и межнейронные взаимодействия определяются генетической специализацией клетки.
Типы нервных сетей. Существуют три генетически детерминированных типа нервных сетей. Чтобы сделать концепцию генетической детерминации нейронных сетей более понятной, давайте уменьшим их число и представим себе, что наша нервная система состоит всего лишь из 9 клеток (см. рис. 33). Это абсурдное упрощение поможет нам проявляется в наличии трех основных типов сетей, которые встречаются повсюду, — иерархические, локальные и дивергентные с одним входом.
Иерархические системы обеспечивают очень точную передачу информации. В результате конвергенции (когда несколько нейронов одного уровня контактируют с меньшим числом нейронов следующего уровня) или дивергенции (когда контакты устанавливаются с большим числом клеток следующего уровня) информация фильтруется и происходит усиление сигналов. Но, подобно любой цепи, иерархическая система не может быть сильнее своего самого слабого звена. Инактивация любого уровня, вызванная ранением, заболеванием, инсультом или опухолью, может вывести из строя всю систему. Конвергенция и дивергенция, однако, оставляют цепям некоторый шанс уцелеть даже при их серьезном повреждении. Если нейроны одного уровня будут частично уничтожены, сохранившиеся клетки смогут все-таки поддерживать функционирование сети.
Дивергентные сети с одним входом. В некоторых нервных сетях имеются скопления или слои нейронов, в которых один нейрон образует выходные связи с очень большим числом других клеток (в таких сетях дивергенция доведена до крайних пределов). Изучение сетей такого типа начато лишь недавно, и единственные места, где они встречаются (насколько нам сейчас известно), — это некоторые части среднего мозга и ствола мозга. Преимущества подобной системы в том, что она может оказывать влияние на множество нейронов сразу и иногда осуществлять связь со всеми иерархическими уровнями, нередко выходя за пределы специфических сенсорных, двигательных и других функциональных объединений.
Сфера воздействия таких сетей не ограничена какой-либо системой с определенными функциями. Дивергирующие пути этих сетей иногда называют неспецифическими и поэтому такие сети могут влиять на самые различные уровни и функции. Они играют большую роль в интеграции многих видов деятельности нервной системы. Кроме того, медиаторы, используемые в дивергентных системах с одним входом, — это медиаторы с «условным» действием: их эффект зависит от условий, в которых он осуществляется. Подобные воздействия весьма важны и для интегративных механизмов. Однако дивергентные сети такого типа составляют лишь небольшую часть всех нервных сетей.
Тема 6. Концевые нервные аппараты и их классификация. Рефлекторная дуга и динамическая поляризация нейронов
Связь нейронов с различными тканями и органами устанавливается при помощи нервных волокон, которые образуют в них концевые нервные аппараты (нервные окончания). Окончания аксонов периферических нервов подразделяют на чувствительные (афферентные) и двигательные (эфферентные).
Приспособления, которые воспринимают раздражения, называются рецепторными аппаратами, или чувствительными нервными окончаниями, а нервы, проводящие возбуждение – чувствительными. Реализация нервных импульсов осуществляется эффекторными аппаратами (двигательными нервным окончаниями), а проведения возбуждения к ним происходит по двигательным нервам.
Концевые нервные аппараты – сложные образования. В их состав входят не только нервные волокна, но и ткани, в которых они оканчиваются. Структура концевых аппаратов разнообразна, меняется в зависимости от условий, в которой они находятся.
Эффекторный аппарат хорошо представлен на двигательной бляшке. Он располагается на поперечнополосатом мышечном волокне в виде разветвления осевого цилиндра мякотного нервного волокна которое теряет миелин. По данным электронной микроскопии, для двигательной бляшки характерно отчетливое разграничение нервной и мышечной частей. В гладких мышцах двигательная иннервация осуществляется безмякотными нервными окончаниями.
Секреторные окончания эффекторных нейронов представлены аксонами, выступающими в Синаптический контакт с железистыми клетками. Концевые разветвления аксона либо подходят вплотную к секреторной клетке, либо глубоко вдавливаются в нее. Нейролемма аксона и плазмалемма секреторной клетки образуют соответственно пресинаптическую и постсинаптическую мембраны, разделенные узкой синаптической щелью.
Холинрецепторы присутствуют также в мембране мышечного волокна вне синапса, но здесь их концентрация на порядок меньше, чем в постсинаптической мембране и обозначаются они как холинрецепторы.
Рецепторные аппараты (рецепторные нервные окончания. Рецепторные (воспринимающие) нервные окончания) у позвоночных представляют собой концевые аппараты дендритов чувствительных нейронов, тела которых располагаются чаше всего в спинальных ганглиях и их аналогах – черепномозговых чувствительных узлах или в периферических вегетативных ганглиях. В зависимости от того, откуда они воспринимают раздражение, различают экстерорецепторы и интерорецепторы. Первые воспринимают раздражения из внешней среды, вторые – из внутренних органов. Кроме того, с учетом специфичности раздражителя различают тактильные, холодовые, тепловые, болевые рецепторы, барорецепторы, хеморецепторы, механорецепторы.
По морфологическим особенностям рецепторные окончания могут быть свободными, располагающимися между клетками иннервируемой ткани, и несвободными, инкапсулированными (заключенными в особые соединительнотканные капсулы).
Свободные нервные окончания – наиболее распространенный тип сенсорных рецепторов. Большинство свободных нервных окончаний – механорецепторы. Распространены в прослойках соединительной ткани внутренних органов, а также в соединительнотканной основе кожи. Свободные нервные окончания эпидермиса расположены в базальном и шиповатом слоях. В области кожи с высокой тактильной чувствительностью (пальцы рук) терминали достигают зернистого слоя. Некоторые окончания в эпидермисе специализированы для регистрации изменений температуры. Свободные нервные окончания имеются и в других органах чувств (слуха, равновесия, вкуса), закладывающихся из эктодермы.
В многослойном эпителии локализованы чувствительные осязательные клетки Меркеля, имеющие округлую или удлиненную форму. Они соединены с эпителиоцитами при помощи десмосом и формируют контакт с нервными терминалями. В клетках Меркеля обнаружены пептиды и нейроспецифические вещества, что свидетельствует об их эндокринной функции. Это позволяет рассматривать их как компонент диффузной нейроэндокринной системы.
К механорецепторам примерно такого же строения относятся луковицеобразные тельца (тельца Гольджи – Маццони), которые расположены в концевой части сухожилий на границе с мышцей, а также в связках капсулы суставов.
В теле человека встречаются концевые колбы (колбы Краузе), которые являются терморецепторами. Они расположены в соединительнотканной основе кожи, слизистых и серозных оболочках. Они также имеют тонкую соединительнотканную капсулу, образующую наружную колбу рецептора. Температурные раздражения воспринимают капсулированные клубочки (тельца Руффини) – крупные рецепторы веретеновидной формы длиной до 2 мм и диаметром около 150 мкм. Они располагаются в соединительной ткани кожи и суставов.
К группе капсулированных нервных окончаний относятся генитальные тельца (тельца Догеля). Они обнаружены в соединительной ткани половых органов, головки полового члена, клитора и других частях тела. По своему строению они напоминают тельца колбы Краузе. Генитальное тельце является механо – и барорецептором, поскольку реагирует на изменение кровяного давления. Из капсулированных механорецепторов кожи птиц наиболее распространены тельца Хербста и тельца Грандри, расположенные в восковице пластинчатоклювых. Тельце Хербста имеют такое же строение, как и тельца Фатер – Пачини. Тельце Грандри мельче телец Хербста и они обладают более тонкой соединительнотканной капсулой. Внутри капсулы находятся две крупные нейроглиальные клетки с крупными овальными ядрами.
Таким образом, инкапсулированные рецепторные окончания всегда состоят из разветвлений осевого цилиндра чувствительного нейрона, оканчивающихся на глиальных клетках, окруженных соединительнотканной капсулой.
Различают простые и сложные рефлексы. Простейший рефлекс выполняется на уровне спинного мозга без участия головного мозга. Такой рефлекс осуществляется при участии трех типов нейронов: чувствительного, вставочного и двигательного.
%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7/%D0%BF%D0%BE%D1%81%D0%BE%D0%B1%D0%B8%D0%B5%20%D0%90%D0%9D%D0%A1%202016%20(%D0%A7%D0%B0%D1%81%D1%82%D1%8C%20I)%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7..files/image031.gif)
%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7/%D0%BF%D0%BE%D1%81%D0%BE%D0%B1%D0%B8%D0%B5%20%D0%90%D0%9D%D0%A1%202016%20(%D0%A7%D0%B0%D1%81%D1%82%D1%8C%20I)%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7..files/image033.gif)
Вставочные нейроны – небольшие мультиполярные клетки с короткими дендритами. Их единственный нейрит проникает в белое вещество, где разделяется на две ветви, одна из которых направляется вверх, а другая – вниз. В выше- и нижележащих отделах спинного мозга они опять заходят в серое вещество и вступают в контакт с двигательными, или моторными, нейронами. Этот тип связующих нейронов характеризуется тем, что их отростки не выходят за пределы спинного мозга и объединяет только его отделы. Кроме таких клеток, в задних рогах и в средней части серого вещества имеется и другой тип связующих нейронов. Их восходящий отросток отличается значительной длиной и поэтому достигает стволовой части головного мозга.
Связующие клетки представляют второй тип нейронов, принимающих участие в осуществлении рефлекса. В них происходит трансформация чувствительного импульса в двигательный. Дальнейший путь этого импульса связан с проводящими волокнами связующих нейронов и наличием в спинном мозге двигательных нейронов. На теле этих нервных клеток оканчиваются отростки вставочных нейронов.
Двигательные (моторные) нейроны расположены в передних рогах серого вещества спинного мозга отдельными группами. Эти – самые крупные клетки спинного мозга. Они являются мультиполярными и отличаются сильно разветвленными дендритами. Аксон этих клеток выходит из спинного мозга по переднему корешку и направляется к мышце. Двигательный импульс по волокнам этих клеток попадает к исполнительному органу, который совершает работу. На этом заканчивается путь чувствительного импульса, который возник в рецепторе.
Белое вещество состоит из волокон, большая часть которых принадлежит к мякотным. Они расположены вдоль спинного мозга и образуют проводящие пути – короткие, объединяющие разные уровни спинного мозга, и длинные, соединяющие спинной мозг с головным.
Тема 7. Оболочки мозга. Желудочки мозга. Ликвор.
Твердая оболочка довольно прочная, она образована плотной волокнистой соединительной тканью. В спинном мозге эта оболочка имеет вид продолговатого мешковидного образования, в полости которого располагаются спинной мозг с корешками спинномозговых нервов, спинномозговые узлы и остальные две оболочки. Наружная поверхность твердой оболочки в спинном мозге отделена от надкостницы жировой клетчаткой и венозным сплетением. Твердая оболочка головного мозга срастается с надкостницей внутренней поверхности костей, в месте прохождения крупных вен, отводящих кровь головного мозга, сращение отсутствует. Твердая оболочка головного мозга состоит из двух пластинок: наружной и внутренней. Наружная пластинка образована плотно упакованными пучками коллагеновых волокон, между которыми проходят кровеносные сосуды, питающие саму оболочку, кости и нервные волокна. Внутренняя пластинка более тонкая, в некоторых участках она глубоко вдается в ткань мозга, образуя синусы. В эти синусы оттекает кровь, которая затем проходит в вены. Серп большого мозга образован внутренней пластинкой твердой оболочки. Твердая мозговая оболочка складкой проникает глубоко между правым и левым полушариями и между затылочным полюсом полушарий и мозжечком.
%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7/%D0%BF%D0%BE%D1%81%D0%BE%D0%B1%D0%B8%D0%B5%20%D0%90%D0%9D%D0%A1%202016%20(%D0%A7%D0%B0%D1%81%D1%82%D1%8C%20I)%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7..files/image035.jpg)
Рис. 12. Оболочки мозга и желудочки
Сосудистая, или мягкая, оболочка прилегает непосредственно к ткани мозга. Эта оболочка образована тонкой рыхлой соединительной тканью, богатой коллагеновыми и эластическими волокнами. В ней проходит богатая сеть кровеносных сосудов. От сосудистой оболочки отходят соединительнотканные волокна, которые вместе с кровеносными сосудами проникают в вещество мозга.
%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7/%D0%BF%D0%BE%D1%81%D0%BE%D0%B1%D0%B8%D0%B5%20%D0%90%D0%9D%D0%A1%202016%20(%D0%A7%D0%B0%D1%81%D1%82%D1%8C%20I)%20%D0%9E%D0%9A%D0%9E%D0%9D%D0%A7..files/image037.jpg)
Рис.13. Желудочки мозга
Общее количество ликвора у человека более или менее постоянно и в норме составляет 120-150 мл. Из этого количества только 20-40 мл находится в желудочках. В норме ликвор прозрачен, бесцветен и его удельный вес колеблется в пределах 1,0, а рН =7,4, т.е. среда является слабо щелочной. В ликворе содержится небольшое количество лимфоцитов, число которых в норме не должно превышать 5 клеток на 1 мл жидкости. В спинномозговой жидкости содержание белков меньше, чем в крови и лимфе. Для сравнения: содержание белков в крови составляет 7-8%, в лимфе – 0,3-05%, а в ликворе всего лишь – 0,02%. Содержание белков в ликворе у детей ниже, чем у взрослых. В ликворе обнаружены в небольшом количестве хлориды, соли кальция, магния, также органические соединения в виде аминокислот, креатина, молочной и мочевой кислот, холестерина и сахара. В норме в спинномозговой жидкости не должны присутствовать ферменты и иммунные тела.
Таким образом, ликвор как бы создает вокруг мозга «водяную подушку» и тем самым предохраняет его механических повреждений при различных резких поворотах и движениях. Кроме того, спинномозговая жидкость способствует удалению из нервной ткани продуктов обмена. Нарушение ее нормальной циркуляции приводит к различным расстройствам центральной нервной системы. При накоплении избыточного количества ликвора развивается гидроцефалия, что угрожает функциональному состоянию мозга.
Спинной мозг питается кровью из глубокой шейной артерии, межреберных, поясничных, подключичных артерий и через позвоночные артерии. Позвоночные артерии проходят в полость черепа и там распадаются две парные передние и задние спинномозговые артерии. Вглубь мозга артерии посылают тонкие веточки. В результате разветвления сонных артерий формируется артериальных круг большого мозга, называемый виллизиевым кругом. Непрерывное снабжение мозга кровью является основным условием его нормальной деятельности. Никакие другие клетки организма так быстро не перестают функционировать, как нервные при недостаточном кровоснабжении. Кратковременное прекращение кровоснабжения мозга приводит к обмороку, что свидетельствует о его высокой чувствительности к постоянному поступлению кислорода и питательных веществ (особенно глюкозы).
3. Анатомия центральной нервной системы: Учебное пособие / О.О. Якименко. – Изд-во М.: Академический проект Трикста, 2009. – 111с.
4. Анатомия центральной нервной системы: Учебное пособие для студентов вузов / Н. В. Воронова, H. M. Климова, А. М. Менджерицкий. — М.: Аспект Пресс, 2005. — 128 с.
6. 4. Мозг, разум, поведение /Под ред. Ф. Блума. – М.: Мир, 1989. – 432с.
7. Шляхтин Г. В. Анатомия и эволюция нервной системы. Саратов, 1984. – 389с.