АНТИГЕНЫ
Антигены (греческий anti- против + gennaö создавать, производить) — любое вещество, которое, поступая в организм парентеральным путем, вызывает ответную специфическую иммунологическую реакцию, проявляющуюся в образовании специфических антител. Попадание антигенов в организм может сопровождаться возникновением состояния толерантности к этому веществу (см. Толерантность иммунологическая) или повышением чувствительности к данному антигену. (см. Аллергия).
Специфическим антигеном может быть определенное молекулярно-гомогенное вещество. Однако антигенные свойства отдельных веществ проявляются и в том случае, если они входят в состав сложных смесей и систем. Поэтому в клинике инфекционных болезней, в лабораторной и эпидемиологической практике термин «антиген» часто используют по отношению к таким сложным системам, как микробные, растительные и животные клетки, тканевые экстракты, биологические жидкости и т. д., имея при этом в виду отдельные содержащиеся в этих системах антигены. Термин «антиген» нередко употребляют и для обозначения веществ, которые, в отличие от полноценных антигенов, не способны самостоятельно стимулировать синтез антител (см.) в организме, но могут специфически реагировать с уже образовавшимися антителами. В иммунологии для определения таких веществ принят специальный термин — гаптены (см.).
По своей природе антигены — высокомолекулярные полимеры естественного происхождения или синтезированные искусственным путем. Свойствами полноценных антигенов обладают белки, полипептиды, полисахариды, а также, вероятно, высокополимерные нуклеиновые кислоты и комплексные соединения этих веществ.
Антигенность определяется не только особенностями химического строения веществ, но зависит также от видовой принадлежности иммунизируемого животного и его генетической конституции (см. Иммуногенетика). Одно и то же вещество, не будучи антигенным по отношению к животным одного вида, вызывает специфическую иммунологическую реакцию при введении особям другого вида. Так, полисахарид декстран не является антигеном для кроликов, а при введении человеку стимулирует синтез специфических антител даже после однократной инъекции. Более того, в пределах одного вида встречаются особи, рефрактерные (не вырабатывающие антитела) и, наоборот, высокочувствительные к данному антигену.
Антигенность как биологическое явление относительна, и для реализации этого свойства необходимо проникновение вещества во внутреннюю среду иммунокомпетентного организма, чувствительного к данному веществу.
Несмотря на огромное число фактов, полученных в ходе химического исследования антигены, иммунология еще не достигла такого уровня, чтобы можно было провести полный перечень тех физико-химических особенностей строения веществ, которые создают необходимую основу для возникновения антигенных свойств. Тем не менее известны некоторые признаки, отличающие антигенные вещества от неантигенных, например, свойствами полноценных антигенов обладают вещества, характеризующиеся, как правило, высоким молекулярным весом — 10 000 и выше.
Функционально активные белки состоят из субъединиц — полипептидных цепей, соединенных друг с другом в единую молекулу дисульфидными или водородными связями. Диссоциация этих связей в ряде случаев приводит к нарушению антигенной специфичности. Так, фермент лактатдегидрогеназа (молекулярный вес 135000) состоит из четырех субъединиц двух генетически различных типов. В отличие от нативного фермента, полипептидные субъединицы в диссоциированном состоянии не только не способны индуцировать синтез специфических антител, но и не реагируют с антисывороткой к нативному ферменту.
Появление антигенной способности с увеличением молекулярного веса веществ характерно не только для белков, но и для полисахаридов. Исследование различных препаратов декстранов с молекулярным весом от 10 000 до 200 000 показало, что стимуляцию антителогенеза у человека вызывают декстраны, молекулярный вес которых не ниже 50 000. Вместе с тем было бы неверно считать, что высокий молекулярный вес является обязательным свойством антигена. Так, сульфированный полистирол — высокомолекулярный полимер — не обладает антигенностью. Нуклеиновые кислоты, несмотря на высокий молекулярный вес, значительно более слабые антигены, чем белки. Сывороточный альбумин и гемоглобин имеют одинаковый молекулярный вес (около 70 000), однако способность индуцировать образование антител у гемоглобина выражена в значительно меньшей степени, чем у альбумина.
Явное исключение из изложенного составляют антигеноактивные вещества, которые характеризуются относительно невысоким молекулярным весом: глюкагон, гормон поджелудочной железы (молекулярный вес 3800) и другие, антигенное действие которых проявляется при иммунизации с адъювантами (см.). Более того, иммунными свойствами могут обладать синтетические полипептиды, молекулярный вес которых равен 4000 и 1200.
Помимо величины молекулы, антигенность вещества определяется также и рядом других его свойств. Одним из необходимых свойств антигенов, как полагают, является жесткость структуры входящих в его состав детерминантных групп. Так, желатина, представляющая собой слабоантигенный белок, денатурированный нагреванием, не обладает фиксированной внутренней структурой; в ее состав входит много глицина, не имеющего в α-положении боковых групп, что обусловливает возможность продольного вращения. Если же ввести в молекулу желатины химической группировки, увеличивающие жесткость ее структуры (тирозин, триптофан, фенилаланин), то она преобразуется в сравнительно сильный антиген. Аналогичного рода данные были получены при изучении антигенных свойств синтетических полипептидов. Повышать жесткость молекул в полисахаридных антигенов могут пиранозные или фуранозные кольца.
Исследование искусственных полипептидов позволило установить роль некоторых аминокислот в проявлении антигенных свойств веществ. При сравнении полипептидов глю58-, тир4-, глю57-, лиз38-, ала5- было показано, что аланин, так же как и тирозин, усиливает иммуногенные свойства полипептида. Установлено снижение влияния глутаминовой кислоты на антигенность полипептида после введения в его состав небольшого количества тирозина.
Свойством антигенов является их способность подвергаться в организме процессам метаболизма. В этой связи интересны данные о роли оптической изомерии аминокислот в определении антигенности вещества. Как оказалось, полипептиды, построенные из L-аминокислот, являются активными стимуляторами антителогенеза, тогда как полипептиды из D-аминокислот способны вызывать образование антител лишь при введении их в малых дозах. В больших дозах D-полипептиды вызывают толерантность.
Антигенная активность веществ, и в частности их способность к индукции синтеза антител, наиболее сильно проявляется, в том случае, если иммунизируемое животное принадлежит к иному, чем источник данного вещества, виду. Общепризнано, что антигенность белков тем выше, чем к более отдаленной таксономической группе относится иммунизируемое животное.
Белки и углеводы крови и внутренних органов обычно не антигенны для организма, в котором они синтезируются, и в то же время антигенны для других особей того же вида. Эта закономерность не распространяется на так наз. забарьерные органы, то есть органы, отделенные от кровотока особыми барьерами (гемато-энцефалический барьер, гемато-тестикулярный барьер и др.), белки которых в норме обычно не поступают в кровь и являются антигенами для собственного организма. В число таких органов входит мозг, хрусталик, паращитовидные железы, семенник.
Толерантность (иммунологическая ареактивность организма к данному антигену) к собственным белкам хорошо объясняется с позиций клонально-селекционной теории иммунитета. Одно из основных положений этой теории утверждает, что «распознавание» собственных белков организма и толерантность к ним связаны с элиминацией в эмбриональном периоде развития всех клонов лимфоидных клеток, способных реагировать против антигена данного организма. С позиций этой теории антигены представляются веществами, несущими в себе признаки чужеродной генетической информации. Следовательно, для того чтобы вещество могло проявить свои антигенные свойства, оно должно отличаться от антигена тканей иммунизируемой особи. Отсюда вытекает, что антигенность вещества зависит и от его специфичности.
С помощью метода комплексных антигенов, то есть антигена, в молекулу которых искусственно введена определенная хим. группировка, было установлено, что антигенная специфичность комплексных антигенов определяется не всей макромолекулой в целом, а свойствами этой группировки — детерминантной группы. При этом оказалось, что специфичность антигенов определяется не только химическим составом детерминантной группы, но и положением ее в антигене, а также пространственным расположением атомов в ней и связанной с этим их стереоизомерией.
В естественных белках антигенная специфичность также определяется небольшой частью ее молекулы. Установлено, что реакцию образования антител против фиброина шелка могут специфически подавлять продукты гидролиза шелка с молекулярным весом, равным всего около 600—1000, причем самыми эффективными в таком подавлении являются глицилаланиновые цепочки длиной в 12 аминокислот (молекулярный вес 900). Из октапептидов наиболее эффективным оказался гли-/гли3-ала3-/тир- с молекулярным весом около 600, который и является главной частью специфической антигенной детерминанты. По данным других исследователей, антигенная специфичность декстрана, синтетических полипептидов (полиаланина, полилизина), миоглобина зависит от небольших реактивных участков с молекулярным весом в пределах 350—990.
Сравнение антигенных свойств у белков с известной последовательностью аминокислотных остатков позволило установить, что для появления новой антигенной специфичности достаточно минимальных изменений в первичной структуре белков. Так, антигенные различия инсулинов у некоторых животных (свиней, крупного рогатого скота, овец, лошадей) обусловлены замещением аминокислотных остатков всего в трех участках полипептидной цепи. Генетические варианты молекул иммуноглобулинов человека [Inv(a+ ) и Inv(a-)] различаются между собой лишь одним аминокислотным остатком в 189-м положении легких цепей, однако этого оказывается достаточно, чтобы они различались как антигены.
Анализ антигенной специфичности синтетических полипептидов показал дальше, что в большой степени их специфичность определяется характером концевых групп. Однако в ряде случаев удавалось отметить существование перекрестных реакций и между полипептидами, концевые группы которых отличались друг от друга. Как было выяснено, такие перекрестные реакции были обусловлены наличием общих аминокислот в других положениях. В последующих опытах было установлено, что антитела могут быть направлены против всего полипептида, состоящего из пяти аминокислот, в целом. Сходные результаты дали и опыты с углеводными гаптенами. Здесь также было выявлено ведущее влияние на специфичность антигена концевых групп, а также показано, что антитела могут быть направлены и против всего гаптена в целом. Самой большой группировкой, которая может реагировать с данным антителом и, следовательно, определять специфичность антигена, являются, очевидно, гексасахариды.
Таким образом, в естественных белках и полисахаридах антигенная специфичность определяется составом и последовательностью аминокислот в полипептидной цепи и моносахаров в полисахариде, особенно их концевыми аминокислотами или моносахарами.
Как известно, вторичная и в конечном счете третичная структура белковой молекулы определяется последовательностью аминокислот. С другой стороны, антигенную специфичность молекулы белка определяют в основном группировки, расположенные на ее поверхности. Поэтому можно утверждать, что антигенная специфичность белка зависит и от его вторичной и, возможно, третичной структуры. Кроме того, приводившиеся выше результаты изучения антигенных свойств лактатдегидрогеназы показывают, что антигенная специфичность высокомолекулярных белков, состоящих из субъединиц, может определяться и их четвертичной структурой.
Образующиеся на поверхности белковой молекулы антигенные детерминанты могут различаться по форме, размерам, по числу и набору входящих в эти детерминанты аминокислот. В результате при иммунизации даже чистым кристаллическим препаратом белка в организме образуются антитела разных типов, неоднородные по своей специфичности. Число антигенных детерминант в молекуле (валентность антигена) варьирует у разных белков в зависимости от размеров молекул: от 5 в молекуле яичного альбумина (молекулярный вес 40 500) до 40 в молекуле тиреоглобулина (мол. вес 650 000). Однако прямой зависимости между валентностью и молекулярным весом антигенов не существует.
Характер взаимодействия антигенных детерминант и остальной части молекулы в определении антигенных свойств вещества пока еще полностью не раскрыт. Тем не менее накопленные факты свидетельствуют о том, что стимуляция иммунологических реакций организма осуществляется реактивными группами молекул антигенов, определяющими его специфичность, то есть детерминантными группами.
Говоря о специфичности природных антигенов в первую очередь имеют в виду их видовую специфичность. Действительно, для особей данного вида присуща антигенная специфичность, не характерная для особей, которые принадлежат к любому другому виду живых существ. Не следует, однако, думать, что имеются какие-то вещества, специально «отвечающие» за антигенную видоспецифичность. Такой видоспецифичностью обладают, очевидно, многие, если не большинство веществ, содержащихся в организме.
Хотя все виды живых существ четко отличаются друг от друга своими видоспецифическими антигенами, степень этого различия может быть неодинаковой. Близкородственные виды характеризуются наличием достаточно сходных видоспецифических антигенов. Видам, далеко отстоящим друг от друга, присущи и резко различающиеся видоспецифические антигены. На основе учета этого явления выросло самостоятельное биологическое направление — иммуносистематнка, использующее метод антигенного анализа для решения сложных таксономических проблем и вопросов эволюционных отношений различных видов микроорганизмов, растений и животных.
Уже в начале нашего века было установлено, что группы различных особей одного и того же вида могут отличаться друг от друга по содержанию антигенов, которые впоследствии получили название изоантигенов. Изоантигены были выявлены в клетках всех изучавшихся видов животных. Однако достаточно полно они изучены лишь у человека. Как оказалось, изоантигенная структура клеток человека исключительно сложна. Только в эритроцитах человека было выявлено более 15 систем изо-антигенов, включающих около 100 антигенов. (см. Группы крови). Подобно тому, как практика переливания крови потребовала развития исследований, приведших к описанию антигенной структуры эритроцитов, наблюдающееся в наши дни повышение интереса клиницистов к пересадке тканей и органов обусловило переход к тщательному изучению антигенного состава других клеток организма. Было установлено, что большинство антигенов, обусловливающих реакцию реципиента против пересаженного органа, содержится в лейкоцитах. Поэтому особое внимание было обращено на изучение антигенов, содержащихся в этих клетках крови. Различные исследователи описали большое количество разнообразных антигенов лейкоцитов. При сопоставлении всех этих антигенов друг с другом оказалось, что большинство их принадлежит к единой системе, получившей название HL-A. Помимо этой системы, пока выявлена еще одна система лейкоцитарных антигенов, генетически независимая от системы HL-A, — система группы 5. Как было установлено, все антигены обеих систем, за исключением, возможно, антигена 9, представлены несколькими аллелями (см.). Было также показано, что эти антигены присутствуют, помимо лейкоцитов, в клетках многих органов и тканей человека, что особенно важно для подбора доноров и реципиентов при пересадке органов в клинике (см. Несовместимость иммунологическая).
Помимо изоантигенов, характерных для эритроцитов и для лейкоцитов, были обнаружены изоантигены, присущие тромбоцитам, лимфоцитам, гранулоцитам, сыворотке крови и др. Поэтому, помимо «общих» изоантигенов, существуют, очевидно, и органоспецифические изоантигены. Вопрос этот, имеющий огромную теоретическую и практическую (при пересадке органов) значимость и в то же время исключительно сложный, в наст, время почти не разработан.
Еще И. И. Мечниковым было установлено, что возможно получение иммунных сывороток, направленных против клеток определенных органов или тканей, — так называемых цитотоксинов. Это открытие легло в основу учения об антигенной органо(ткане)специфичности. Существование органоспецифических антигенов было показано практически во всех органах. Были получены данные о том, что в ряде органов существует два типа органоспецифических антигенов, которые встречаются в одноименных органах представителей разных видов живых существ, и антигены, характеризующие органы только представителей данного вида.
В настоящее время для большинства органов (печень, почка, хрусталик глаза и др.) исследованы главным образом водорастворимые органоспецифические антигены, которые представляют собой более или менее сложные системы белков. Что же касается органоспецифических антигенов, не переходящих в экстракты, то о них имеются лишь единичные отрывочные данные. В самое последнее время были обнаружены антигены, общие для почки, печени, селезенки, сердца, но отсутствующие в сыворотке крови. Некоторые исследователи выделяют их в новую группу — межорганных антигенов.
К группе описанных антигенов примыкают выделяемые некоторыми исследователями так называемые органоидные антигены, характеризующие антигенную специфичность клеточных ядер, митохондрий, рибосом и т. д.
В последние годы установлено существование антигенов, характерных для организмов, их органов или тканей, находящихся на определенных стадиях индивидуального развития. Эти антигены получили название стадиоспецифических антигенов.
Для патологии существенное значение имело обнаружение так называемых патологических антигенов, возникающих в результате патологических процессов. К ним относятся «раковые», «ожоговые», «лучевые» и другие антигены, образующиеся в патологически измененных тканях. Доказано появление новых антигенов (трансплантационных, комплементфиксирующих и поверхностных) в клетках опухолей, индуцированных вирусами.
Антигенная специфичность веществ клеток и тканей отражает существенные особенности их строения, функции и физиологического состояния. Вскрытие причин антигенного действия веществ, анализ их свойств, выяснение химических основ антигенной специфичности веществ — все эти вопросы являются одними из основных вопросов современной иммунохимии. Вместе с тем исследование свойств природных антигенов в наст, время не ограничивается рамками собственно иммунохимии и инфекционной иммунологии и служит для решения многих вопросов, имеющих общебиологическое значение, и, в частности, вопросов эволюции животного и растительного мира.
Анализ антигенных свойств вирусов, бактерий, клеток и тканей многоклеточных организмов показал исключительную сложность их антигенного строения. Наряду с антигенами, свойственными группам особей или всем особям, относящимся к одному виду (видовые, групповые антигены бактерий, изоантигены), в тканях животных присутствуют антигены, распространенные более или менее широко у представителей других видов. Важное значение имело установление того факта, что в определенной степени общие антигены, за исключением гетерогенных антигенов типа антигенов Форссмана, отражают генеалогические связи между видами, у которых они встречаются.
Разные ткани организма отличаются по степени межвидового сходства их антигенов. Сыворотка крови, печень, селезенка и некоторые другие внутренние органы содержат преимущественно антигены с сильно выраженной видовой специфичностью. Напротив, антигены мышц, семенников, мозга, хрусталика по своей специфичности мало отличаются от антигенов гомологичных органов и тканей у представителей разных видов млекопитающих и даже в целом у позвоночных. Это объясняется сходством химического строения и свойств соответствующих белков, несущих одинаковую функцию. Очевидно, в процессе эволюции, на каком-то ее этапе, было достигнуто исключительно полное приспособление структуры таких белков для выполнения функций, имеющих жизненно важное значение, в результате чего все последующие мутации, нарушавшие это соответствие, элиминировались естественным отбором. Как правило, такими общими антигенами являются белки, характеризующиеся крайне слабой антигенностью (гемо-глобины, инсулины, карбомилсинтетаза), или белки тканей, анатомически изолированных от лимфоидной системы организма (белки хрусталика).
Другой причиной существования общих антигенов у представителей разных видов, особенно у видов, филогенетически далеко отстоящих друг от друга, является антигенная адаптация видов-паразитов к антигенам хозяина, отражающая экологические связи между видами.
Некоторые антигены высокоорганизованных животных и, в частности, человека несут защитную функцию в поддержании генетического постоянства внутренней среды организма. Установлено, что антигены системы AB0 (см. Группы крови) присутствуют не только в тканях, но и в виде водорастворимых антигенов в биологических жидкостях и секретах. Объясняя возможное значение явления секреции антигенов, П. Н. Косяков предположил, что антигены AB0 в слюне и в верхних отделах желудочно-кишечного тракта играют защитную роль, нейтрализуя содержащиеся в пище гемагглютинины животного или растительного (лектины) происхождения. Групповые антигены семенной жидкости предохраняют мужские половые клетки от воздействия изоантител, находящихся в женских половых путях в момент оплодотворения.
В явлениях групповой несовместимости материнского организма и плода изоантигены (системы AB0 и др.) последнего, находясь в околоплодной жидкости, амнионе и хорионе, играют защитную роль, связывая антитела матери, проникающие через плаценту, и не «допуская» их к тканям плода.
В последние годы некоторыми исследователями выдвигается положение о возможной морфогенетической роли антигенов в эмбриогенезе (см. Иммунология эмбриогенеза).
Биологическое значение антигенов безусловно не ограничивается их участием в рассмотренных выше явлениях. Так, например, в последнее время интенсивно исследуется вопрос о связи изоантигенов крови с предрасположенностью лиц, дифференцированных по этим антигенам, к некоторым видам заболеваний.
Библиография: Актуальные вопросы иммунологии, под ред. Л. А. Зильбера и П. А. Вершиловой, с. 312, М.. 1964, библиогр.; Бойд У. Основы иммунологии, пер. с англ., М., 1969, библиогр.; Гауровиц Ф. Иммунохимия и биосинтез антител, пер. с англ., М., 1969, библиогр.; Зильбер Л. А. и Абелев Г. И. Вирусология и иммунология рака, М., 1962, библиогр.; Косяков П. Н. Иммунология изоантигенов и изоантител, М., 1965, библиогр.; Петров Р. В. Иммунология острого лучевого поражения, М., 1962, библиогр.; Туманов А. К. Сывороточные системы крови, М., 1968, библиогр.; Эфроимсон В. П. Введение в медицинскую генетику, М., 1968, библиогр.; Andersson В. Interaction between immunocompetent cells and antigen, Stockholm, 1972, bibliogr.; Immunological tolerance to microbial antigens, ed. by H. Friedman, N. Y., 1971. bibliogr.; Kissmeyer-Nielsen F. a. Thorsby E. Human transplantation antigens, Copenhagen, 1970; Strong and weak histocompatibility antigens, Copenhagen, 1970, bibli-ogr.; Surface antigens on nucleated cells, Copenhagen, 1971, bibliogr.
О. E. Вязов, В. М. Барабанов.
Антитело: лучший способ распознать чужого
Универсальная структура антитела позволяет распознавать не только чужеродные, но и собственные молекулы организма, а также передавать различные сигналы между клетками
Автор
Редакторы
Процесс связывания антигена с антителом состоит из множества этапов. Организму нужно создать разнообразные антитела, научить их отличать свои антигены от чужих, отобрать лучшие варианты и заставить клетки их массово производить. И это только начало иммунного ответа: связывание с антигеном влечет за собой длинную цепь молекулярных и клеточных взаимодействий, приводящих к уничтожению врага. Мы попробуем описать сложную жизнь антител в организме, поговорим о разных видах антител (не только у человека) и о том, как люди научились использовать оба свойства антител — узнавать чужого и запускать иммунный ответ — в научных и медицинских целях. Эта статья — вторая в цикле работ, посвященных терапевтическим антителам.
Терапевтические антитела
Спецпроект об антителах, истории их изучения, методах работы с ними, а также о применении антител в современной медицине и биотехнологии.
Партнер спецпроекта — Департамент вычислительной биологии одной из крупнейших российских биотехнологических компаний — BIOCAD. BIOCAD заслужил серьезные позиции на мировом фармацевтическом рынке благодаря выпуску лекарственных препаратов на основе антител.
Эта статья — вторая в цикле работ о терапевтических антителах. Первый текст был посвящен истории изучения антител и введения их в медицинскую практику [1]. По результатам сотни лет исследований мы знаем, что антитела — это молекулы, которые производятся В-лимфоцитами (В-клетками). Они обладают постоянной (константной) и изменяющейся (вариабельной) частями и умеют, с одной стороны, распознавать антиген, а с другой — активировать клетки иммунной системы и запускать иммунный ответ. Попробуем подробнее разобраться, откуда появляются антитела и как они живут в организме до и после встречи с антигеном.
Как узнать чужого
Иммунная система — это система охраны организма от внешних вторжений, например, паразитов (о том, как она работает, подробно рассказано в статье «Иммунитет: борьба с чужими и. своими» [2]). Для того, чтобы быстро обнаружить нарушителя порядка, необходимы дозорные — молекулы, которые смогут его распознать и отличить от собственных клеток. Если у группы паразитов есть какой-нибудь общий и неизменный отличительный признак, это существенно упрощает задачу. Примером могут служить бактерии, чьи жгутики по строению принципиально отличаются от эукариотических: они состоят из белка флагеллина, который в нашем организме не встречается. В таком случае нам достаточно иметь всего одну молекулу, распознающую флагеллин, и она будет сигнализировать о присутствии любой бактерии со жгутиком.
Значения терминов, выделенных полужирным шрифтом, можно посмотреть в «Словарике» в конце статьи.
Иммунная система человека делится на две большие ветви — врожденный и приобретенный (адаптивный) иммунитеты, — за изучение которых в 2011 году была вручена Нобелевская премия [3]. Врожденный иммунитет способен защищать организм сразу же, как только в того проник патоген. Приобретенный же иммунный ответ разворачивается намного дольше, однако действует более изощренно и, кроме того, быстро активируется при повторном контакте с врагом. Врожденный иммунитет можно сравнить со стремительным авангардом, который в случае серьезной угрозы зовет на помощь основные силы организма — приобретенный иммунитет. — Ред.
Но что делать, если в организм попал паразит без РАМР? Или даже не весь паразит целиком, а отдельные его части или продукты обмена? Нужно создать систему, которая не будет заточена на узнавание набора молекул, а сможет точечно опознать практически любую незнакомую молекулу и запустить иммунные реакции. Такую молекулу, которая может вызвать в организме иммунный ответ, называют антигеном.
Антигеном может стать далеко не любая молекула. Для того чтобы иммунитет на нее отреагировал, должны выполняться два важных условия. Первое — чужеродность. Молекула должна быть незнакомой, то есть непохожей на собственные молекулы организма. Пусть у нас есть белок — последовательность аминокислот. В опухолевой клетке белок мутирует, и аминокислоты заменяются на другие. Чем больше таких аминокислотных отличий от исходной молекулы, тем сильнее иммунный ответ, который вызовет такой белок. А если это не собственный белок, а чужой, не имеющий аналогов в организме, то ответ будет максимальным.
Аминокислоты, распознаваемые антителом, могут находиться как друг за другом, так и в разных участках белка. Последовательно расположенные аминокислоты образуют линейный антиген. Если же аминокислоты расположены далеко, то они могут сближаться, так как белки находятся, как правило, в свернутом состоянии (так называемые вторичная и третичная структуры). При этом возникает конформационный антиген (рис. 1).
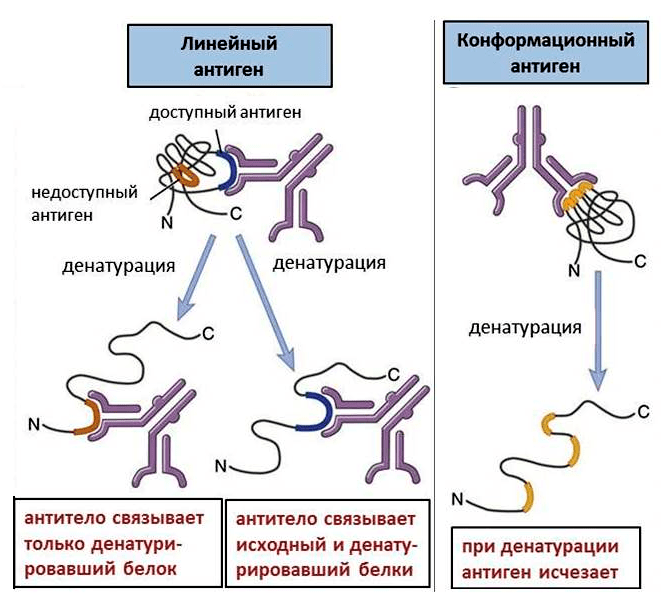
Рисунок 1. Линейные и конформационные антигены. Конформационные антигены доступны для связывания только в интактном белке; при денатурации они исчезают. Линейные антигены присутствуют в любом белке, но денатурация делает некоторые из них доступными.
Чужеродность зависит еще и от того, с какими собственными молекулами иммунная система встречалась раньше [2]. Если, например, какой-нибудь белок находится в глазном яблоке, где нет кровеносных сосудов, то у иммунной системы нет возможности с ним познакомиться и узнать, что он свой. При повреждении глазного яблока белок может попасть в кровоток, и тогда иммунная система распознает его как чужой. Собственные антигены организма называют аутоантигенами в противоположность чужим — аллоантигенам. Когда иммунная система реагирует на аутоантигены, развиваются аутоиммунные заболевания (На «Биомолекуле» есть спецпроект, посвященный тому, почему такие заболевания возникают, и как их лечить).
Второе условие, необходимое, чтобы молекула стала антигеном, — иммуногенность (рис. 2). Иными словами — иммунная система должна иметь возможность встретиться с молекулой и ее распознать. Поэтому, например, антигенами не могут быть мел или масло — они просто не растворяются в воде. Еще антигеном не может быть очень маленькая молекула, например, отдельно взятая аминокислота — ее сложно уловить в растворе, она не свяжется прочно с антителом. Поэтому, например, для белков минимальный размер антигена — от 7 аминокислот. Но многие вещества (например, отдельные металлы) могут стать антигенами, если связаны с белком. В таком случае белок называют носителем: он «отвечает» за то, чтобы иммунная система смогла встретиться с новым веществом, а сам антиген — гаптеном: он «отвечает» за специфичность связывания. Используя носители, можно вызвать иммунный ответ на самые разные вещества, например, анилин [4] или никель [5].
Напротив, большие молекулы хорошо вызывают иммунный ответ: чем длиннее молекула, тем больше в ней участков, которые можно узнать — эпитопов. Также иммуногенность зависит от жесткости структуры — если молекула будет постоянно изменять структуру, то не получится уловить конкретный эпитоп. Поэтому, например, желатин (длинные нити) практически не вызывает иммунный ответ, если его искусственно не стабилизировать. Наконец, чтобы обладать иммуногенностью, антиген должен напоминать по структуре собственные молекулы организма. Это связано с тем, что клетки иммунной системы периодически поглощают антигены, расщепляют их и «демонстрируют» друг другу (см. ниже). А чтобы антиген было легко переварить, он должен быть похож по структуре на собственные молекулы или на молекулы, которыми питается организм, — для них в клетках есть расщепляющие ферменты. Таким образом, в большинстве случаев антигены — это белки или углеводы, в то время как остальные молекулы (например, полиэтилен или другие небиологические полимеры) вызывают иммунный ответ реже и слабее.
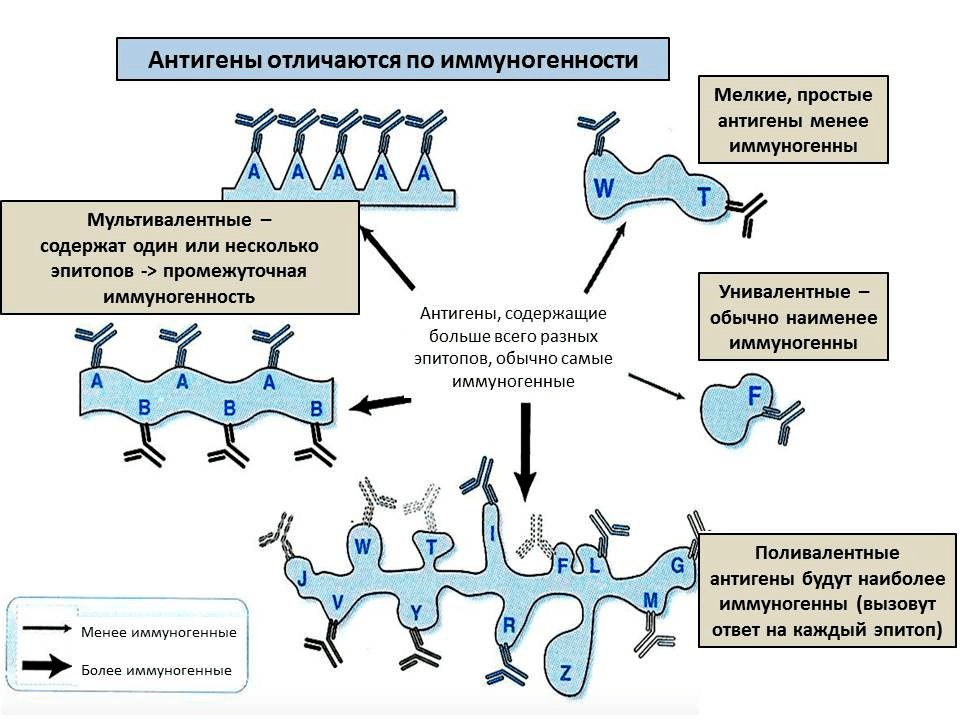
Рисунок 2. От чего зависит иммуногенность антигена? Каждое антитело специфично к одному эпитопу и может реагировать на близкие к нему по строению эпитопы. Чем больше эпитопов на молекуле и чем выше их разнообразие, тем сильнее иммунный ответ на антиген.
Итак, чтобы сигнализировать о патогене, необходима молекула, распознающая небольшие отличия в белках и углеводах и активирующая клетки иммунной системы. Такими молекулами служат антитела, или иммуноглобулины (Ig).
Как устроено антитело
Антитела — это растворимые белки, которые производят В-лимфоциты. У незрелых или покоящихся В-клеток на мембране закреплен предшественник антитела — В-клеточный рецептор, посредством которого В-клетка определяет присутствие антигена. И антитела, и В-клеточные рецепторы построены по одному принципу (рис. 3). Это белки, которые состоят из четырех аминокислотных последовательностей (цепей): двух тяжелых (H-цепи) и двух легких (L-цепи), прочно соединенных дисульфидными связями попарно и между парами. Два конца тяжелых цепей образуют константную часть; она не создает разнообразия и бывает всего нескольких типов. Антитела с одинаковой константной частью составляют один изотип или класс. Самые распространенные изотипы: IgG, IgM, IgD, IgA и IgE. Константная часть определяет, как антитело будет взаимодействовать с клетками или другими антителами:
С другого конца антител находится вариабельная часть, обеспечивающая их разнообразие. Концы легких и тяжелых цепей образуют две одинаковые ямки — антигенсвязывающие участки. Антитела, несущие одинаковые вариабельные части, составляют один идиотип (и могут связывать один и тот же антиген). Антитела разных идиотипов связывают разные антигены благодаря различию в форме и зарядах антигенсвязывающих участков. Можно представить себе антитело как туфельку, которая должна найти свою Золушку. Собственно отверстие туфельки — это антигенсвязывающий участок, куда помещается нога (антиген) претендентки, а константная часть — каблук, за который ее может ухватить принц (иммунная клетка).
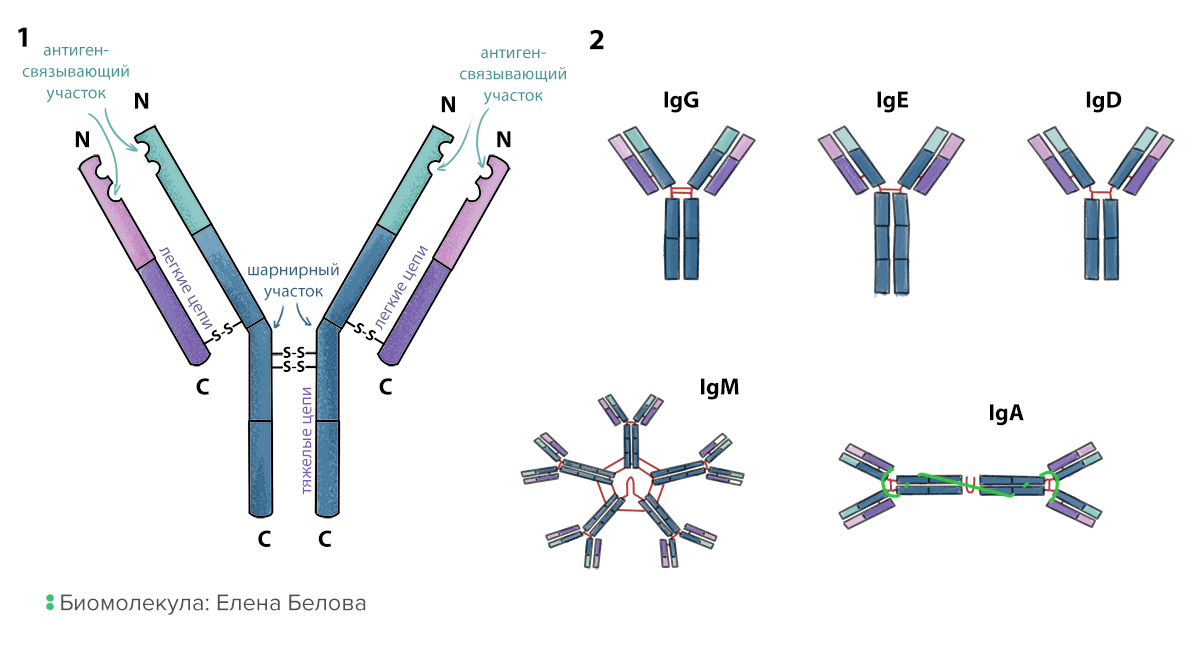
Рисунок 3. Строение и типы антител. 1. Схема строения антитела. Две тяжелые цепи находятся внутри молекулы, две легкие — снаружи. Все они сшиты друг с другом дисульфидными мостиками (S-S). 2. Изотипы антител. Они определяются типом константной части. Некоторые изотипы могут образовывать димеры (IgA) и пентамеры (IgM) с помощью соединительной цепи (joining chain).
Антитела производят В-лимфоциты (или В-клетки). Каждый В-лимфоцит синтезирует свой идиотип антитела. Всего в нашем организме существует около миллиона типов В-клеток. У каждого человека этот миллион немного разный: это зависит не только от отличий в генах иммуноглобулинов, но и от того, как антитело формируется и какие антитела отбирает для себя организм (подробнее об этом можно прочитать в статье «Анализ индивидуальных репертуаров Т-клеточных рецепторов» [6]). Откуда берется такое разнообразие антител, учитывая, что ДНК во всех клетках организма изначально одинаковая, а генов иммуноглобулинов — ограниченное количество?
Как добиться разнообразия
В геноме человека содержится несколько кластеров генов, кодирующих иммуноглобулины, — по одному кластеру для каждой цепи (тяжелой и легкой). Цепи состоят из следующих частей: константная (неизменная), вариабельная (V — variable) и связывающая их (J — joining). В тяжелой цепи между участками V и J есть дополнительная, «разнообразная» часть (D — diversity). Кластер генов содержит множество сегментов — вариантов V, D и J-частей. Из них каждая молодая В-клетка случайным образом отбирает себе по одному, создавая уникальную последовательность антигенсвязывающих участков. Этот процесс называют рекомбинацией генов иммуноглобулинов, или V(D)J-рекомбинацией (рис. 4).
Происходит это следующим образом. Все сегменты окружены специальными сигнальными последовательностями нуклеотидов (recombination signal sequences, RSS). Между каждыми двумя V-сегментами расположен набор из 7, 23 и 9 нуклеотидов. Между каждыми двумя J-сегментами — из 9, 12 и 7. Рекомбинация происходит с участием ферментов RAG: они связываются случайным образом с одной из RSS между V-сегментами и с другой — между J-сегментами. Последовательности из 7 и 9 нуклеотидов комплементарны друг другу, поэтому образуется шпилька, в середине которой — все промежуточные, ненужные V- и J-сегменты. Ферменты катализируют образование разрывов в ДНК, вырезая шпильку. При этом два сегмента до и после шпильки соединяются. Таким образом, клетка отрезает «ненужные» части от своей ДНК, оставляя последовательность иммуноглобулина из константной части и по одному V-, D- (в тяжелой цепи) и J-сегменту. Это первый этап, на котором возникает разнообразие. Учитывая, что V- и J-сегментов содержится в геноме по нескольку десятков (а в тяжелой цепи добавляется еще и около десяти D-сегментов), в результате рекомбинации можно образовать тысячи вариантов цепей.
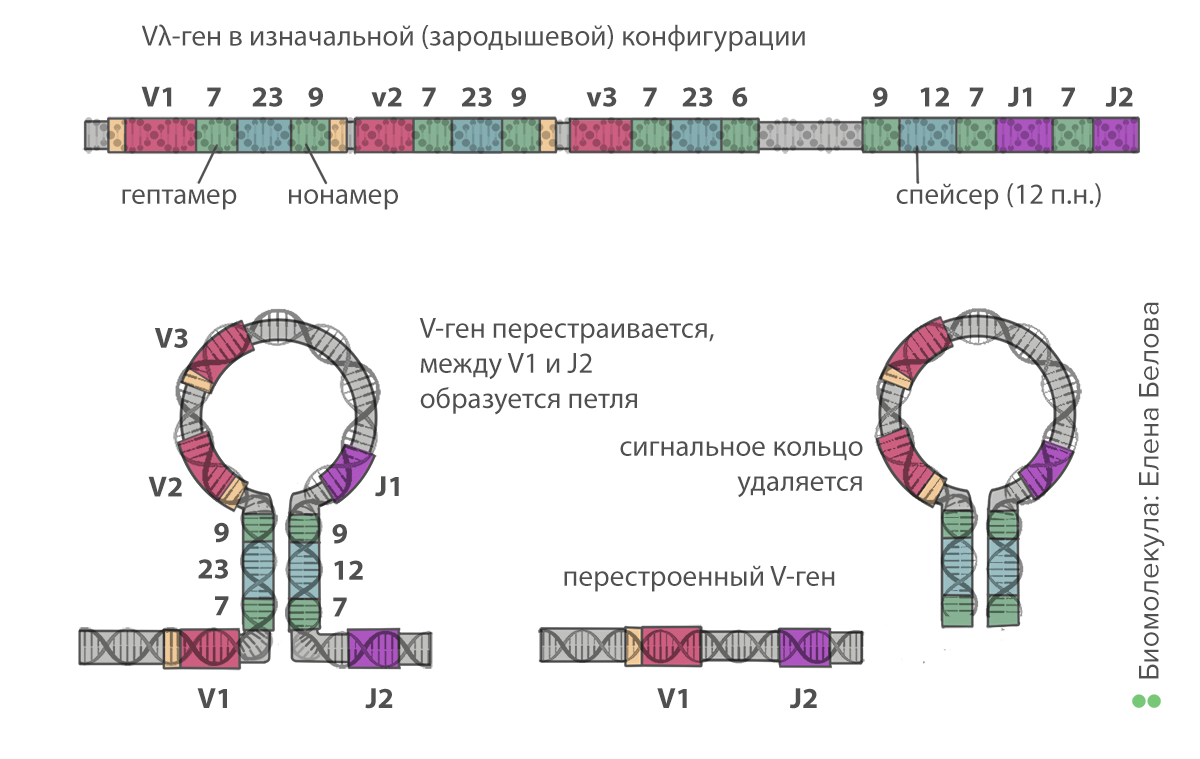
Рисунок 4. Рекомбинация генов иммуноглобулинов, первый этап. Сегменты в каждой группе обозначены цифрами — V1, V2, V3, J1, J2 и так далее. Нонамер (9 нуклеотидов) и гептамер (7 нуклеотидов) — сигнальные последовательности нуклеотидов, комплементарные друг другу. Их связывание друг с другом позволяет образовать кольцо. Таким образом остаются соединены только один из V- и один из J-сегментов.
иллюстрация Елены Беловой по книге Ярилина А.А. Иммунология («ГЭОТАР-Медиа», 2010)
В-клетка может переключаться с одного изотипа антитела на другой, меняя его константную часть. Происходит это с помощью механизма, похожего на рекомбинацию. Гены, кодирующие константную часть, расположены друг за другом (M, D, G, A, E) и разделены S-последовательностями (от switch — переключение). Следовательно, можно соединить две S-последовательности с образованием шпильки и вырезать то, что между ними. Таким образом, переключение изотипов оказывается необратимым: если один раз вырезать участок, кодирующий, например, константную цепь типа G, вернуть его обратно не получится.
У развивающейся В-клетки иммуноглобулин изначально относится к классу М и прикреплен к мембране в составе B-клеточного рецептора (BCR). Этот рецептор нужен, чтобы сортировать В-клетки и избирательно активировать только те, чье антитело подходит для борьбы с конкретным патогеном. Только после активации клетка начинает производить антитело и выбрасывать его в окружающую среду. Таким образом, каждая В-клетка может производить и антитела, и В-клеточные рецепторы одинаковой специфичности. Это две молекулы, очень похожие по строению, но разные по функциям, которые не стоит путать. Кроме того, в организме человека есть и еще одна молекула, похожая по строению на антитело — Т-клеточный рецептор. Она находится на поверхности Т-лимфоцитов и тоже нужна для распознавания антигена. Но это уже совсем другая история — о взаимодействии Т-лимфоцитов с клетками организма [7].
Как работает антитело
Допустим, перед нами уже активированная В-клетка, которая производит и выделяет антитела. Куда им отправиться на поиски антигена? Здесь возможно несколько вариантов.
Растворимые антигены можно встретить свободно плавающими в крови. Это могут быть частички покровов патогена, продукты его обмена веществ или испорченные белки собственного организма — например, мутировавшие, как в случае раковых клеток, или неправильно свернутые. Последствия встречи антитела с растворимым антигеном могут быть разными.
Поверхностные антигены ждут антитела на мембранах клеток. Это могут быть клетки бактерий или собственные клетки человека, например, пораженные вирусом. В случае бактерий антитела тоже могут блокировать работу антигена (например, если прилипнут к белкам бактериального жгутика, — тогда бактерия не сможет передвигаться) или служить опсонином. На антитело, связанное с клеткой, снова реагирует комплемент — в результате каскада реакций в мембране образуются сквозные каналы, и клетку буквально превращают в решето. Кроме того, «черная метка» служит сигналом для иммунных клеток, запуская фагоцитоз или выделение токсичных веществ (рис. 5) [8].
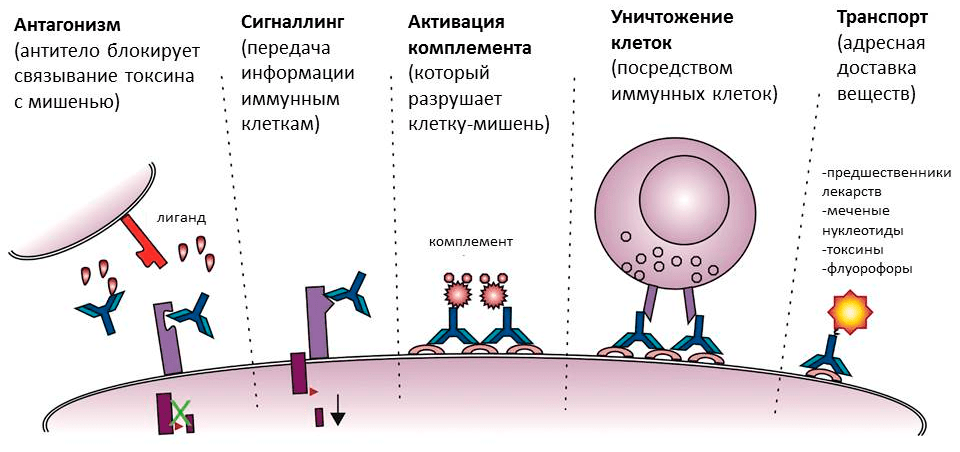
Рисунок 5. Разнообразие функций антител. Помимо прямой — связывания с мишенью, — антитела обладают также набором других функций. Они активируют комплемент и иммунные клетки, направляя их действие на мишень, а в медицине и молекулярной биологии могут использоваться как специфические транспортеры веществ.
Презентация антигена
Кроме того, в организме есть клетки-шпионы, их называют антигенпрезентирующими клетками. К ним относятся специализированные клетки в тканях (дендритные клетки), а также макрофаги и В-клетки. Они несут на поверхности комплекс МНС II типа (МНС II), с которым связаны фрагменты белков, захваченных клеткой из окружающей среды. Клетки-шпионы постоянно поглощают вещества извне, пропускают их через протеасому и так же выставляют на поверхность. Это уже не опись имущества, а настоящий донос — «а я видел, тут мимо проплывал. ». Сами по себе антитела не узнают антигены, связанные с МНС. Зато их отлично узнают родственные антителам Т-клеточные рецепторы, что позволяет им атаковать клетки, демонстрирующие необычные белки (на МНС I) и активироваться при «доносе» о необычных белках (на МНС II) (рис. 6) [9].
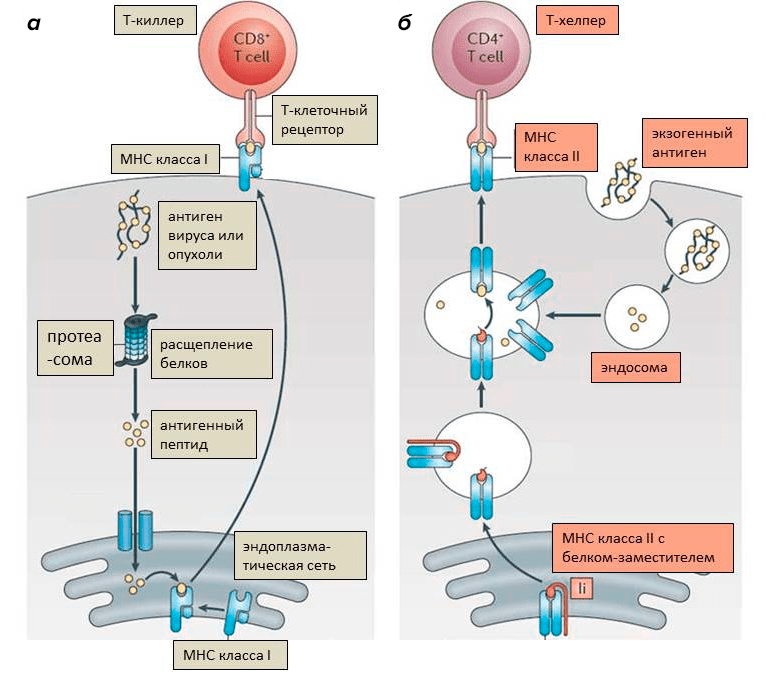
Рисунок 6. Функционирование комплексов МНС. а — Комплекс МНС I связывается с расщепленными фрагментами внутриклеточных белков. Он презентирует антиген Т-киллерным лимфоцитам. Если антиген Т-киллеру не знаком, значит, в клетке находятся вирусные или мутировавшие (опухолевые) белки, и клетка подлежит уничтожению. б — Комплекс МНС II соединяется с фрагментами белков, поглощенных извне (экзогенных). Он презентирует антиген Т-хелперным лимфоцитам. Когда они узнают «свой» антиген, они активируются и могут, в свою очередь, активировать Т-киллера или В-клетку.
Жизнь антител в организме
Теперь соберем весь пазл вместе. Проследим за жизнью В-лимфоцитов и их антител с самого начала. Молодые В-клетки развиваются в красном костном мозге. Там, под действием окружающих клеток соединительной ткани, каждой В-клетке предстоит сформировать свое собственное антитело.
Как мы уже писали выше, В-клетки умеют создавать множество уникальных генов иммуноглобулинов. Но не все эти гены будут рабочими — где-то возникнут поломки из-за многочисленных разрезов и перестроек, где-то итоговый белок не сможет принять правильную структуру. Следующая веха в жизни В-клетки — проверка профпригодности антител. Сначала В-клетка перестраивает ген тяжелой цепи на одной из двух хромосом. Потом синтезирует соответствующие белковые цепи, собирает из них псевдоантитело (вместо легких цепей в нем суррогатные цепи) и выставляет его на мембрану. Если все прошло успешно, то антитело с поверхности клетки посылает сигнал, блокирующий дальнейшую перестройку тяжелой цепи и программу апоптоза — механизма клеточного самоубийства, при котором клетка расщепляет себя изнутри. Если псевдоантитело по каким-то причинам собрать не удалось, запускается перестройка генов на второй хромосоме. Если и со второй попытки не удается сделать рабочую цепь, включается программа апоптоза, и клетка гибнет. Если же тяжелая цепь собрана правильно, у клетки есть две попытки сформировать работающую легкую цепь. Как только иммуноглобулин полностью собран, он появляется на мембране В-лимфоцита, и перестройка генов заканчивается. Теперь у клетки появилось свое уникальное антитело и своя специфичность.
После того, как В-лимфоцит собрал свое антитело, он проходит через строгий отбор (селекцию). Цель его — уничтожить клетки, которые реагируют на собственные антигены организма и могут вызвать аутоиммунную реакцию. А таких среди молодых В-лимфоцитов до 75%! Селекция устроена следующим образом: вокруг В-клеток в красном костном мозге сидят поддерживающие клетки, каждая из которых «показывает» В-клеточному рецептору свои антигены. Каждое связывание В-клеточного рецептора с антигеном посылает в клетку сигнал, стимулирующий апоптоз. В то же время, на поверхности В-клеток есть рецепторы к белкам BAFF и APRIL, которые тоже находятся на поддерживающих клетках. Через рецепторы к BAFF и APRIL поступает сигнал, запрещающий апоптоз. Но этих рецепторов довольно мало. Поэтому, если В-клетка хорошо связывает антигены на окружающих клетках, то проапоптотических сигналов становится больше, чем противоапоптотических, и клетка погибает. А если она связывает антигены плохо или не связывает вообще, то сигналов от BAFF и APRIL рецепторов достаточно для ее выживания. При условии, что селекция работает нормально, из всего разнообразия В-клеток и их антител выживают только те, которые узнают что-то, отличное от собственных молекул организма (рис. 7).
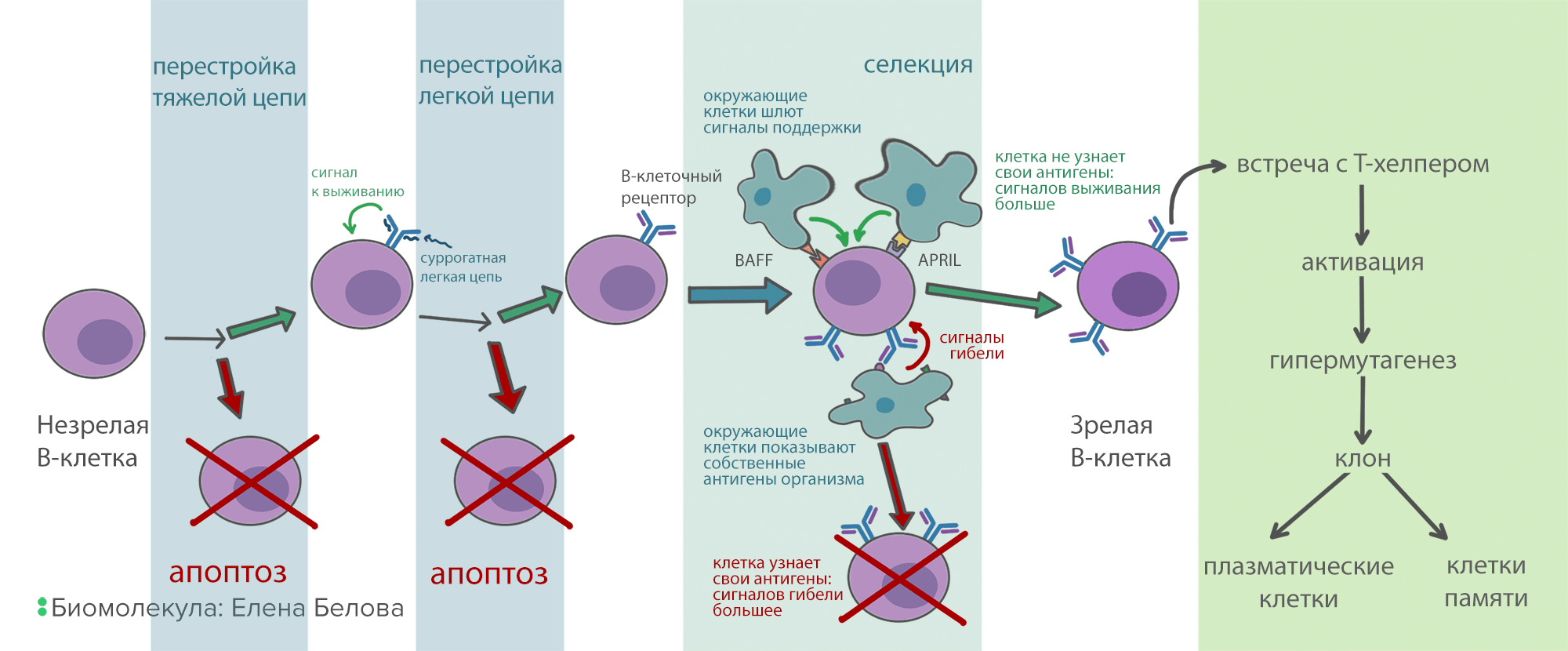
Рисунок 7. Схема развития В-клеток. Прежде чем зрелая В-клетка будет готова синтезировать антитела, она проходит многоэтапную селекцию. Это нужно, чтобы отсеять нерабочие и опасные для организма варианты антител.
Молодая В-клетка плавает с током крови по организму до тех пор, пока не встретит свой антиген. В-клеточный рецептор (BCR) связывается с антигеном, но этого недостаточно для активации. Нужно, чтобы иммунная система официально подтвердила — да, этот антиген действительно опасен. Поэтому В-клетка поглощает комплекс ВCR—антиген, разрезает антиген на части и выставляет их на поверхность в составе комплекса МНС-II. Тем самым она сигнализирует о том, что может создать антитело к вот такому антигену.
В это же время по организму путешествуют Т-хелперные лимфоциты, клетки-помощники. У них есть Т-клеточный рецептор, тоже специфичный к конкретному антигену. Для выполнения своих функций Т-хелперам надо активироваться. Это происходит при встрече с антигенпрезентирующей клеткой, несущей в составе MHC-II антиген, подходящий к Т-клеточному рецептору. Антигенпрезентирующими клетками могут быть как «профессионалы» (дендритные клетки), так и сами В-клетки. Проконтактировав с ними, Т-хелпер получает сигнал тревоги и активируется. Теперь он «знает», что в организме есть такой антиген, и способен активировать В-клетку, если они встретятся. Почему Т-хелпер не может сразу в ответ активировать В-лимфоцит, если он сам выступает в роли антигенпрезентирующей клетки? Контакт с антигенпрезентирующей клеткой протекает очень быстро, а Т-хелперу требуется время, чтобы синтезировать необходимые для активации вещества, поэтому он просто не успевает это сделать и уходит в поисках других подходящих В-клеток. Долгожданная встреча обычно происходит в лимфатических узлах или более крупных лимфоидных органах — красном костном мозге и селезенке. В-клетка показывает с помощью МНС-II антиген, который она способна узнать. Т-хелпер связывается с этим антигеном, «подтверждая», что это тот самый опасный антиген, который уже встречался в организме, и активирует В-клетку (рис. 8) [10].
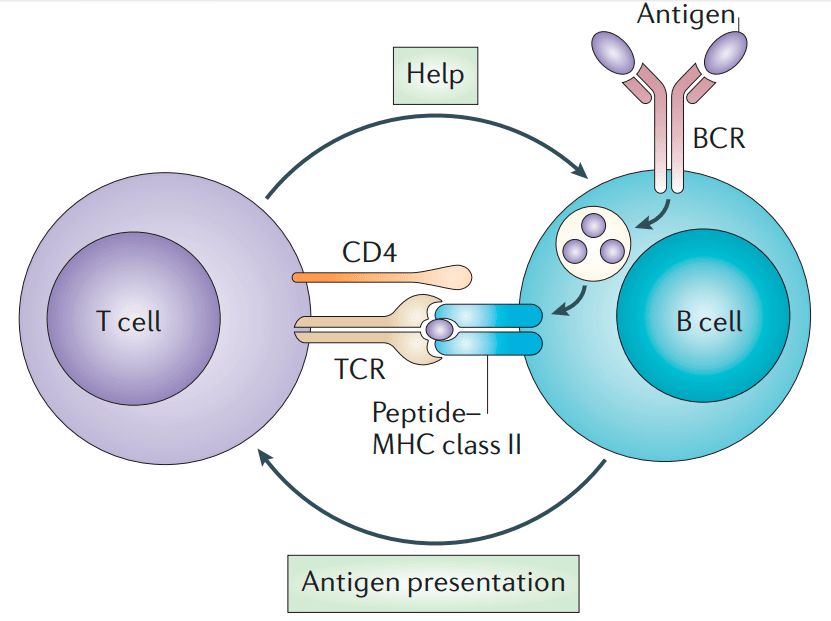
Рисунок 8. Схема встречи В-клетки и активированного Т-хелпера. В-клетка связывает антиген с помощью рецептора (BCR), поглощает его и выставляет в составе МНС-II. Активированный Т-хелпер связывается с МНС с помощью молекулы CD4 и узнаёт антиген своим Т-клеточным рецептором (TCR). Затем Т-хелпер выделяет интерлейкины (IL2/4/5), которые связываются с соответствующими рецепторами (ILR) на В-клетке и активируют ее.
Затем происходит процесс усовершенствования антител — соматический гипермутагенез. В-лимфоцит делится, образуя клон — группу клеток, производящих одинаковые антитела. Клетки клона получают шанс сделать антитело, которое еще лучше связывается с их антигеном, чем предыдущая его версия. При этом клетки случайным образом заменяют нуклеотиды в вариабельных частях гена иммуноглобулина, создавая разнообразные вариации на тему исходного антитела, точнее, его антигенсвязывающей части. Те из них, которые будут распознавать антиген лучше всего, получат сигнал к делению, и образуют окончательный В-клеточный клон. Все клетки клона способны производить один и тот же идиотип (с одинаковой вариабельной частью), но могут переключаться между изотипами (изменять константные части) в зависимости от условий. Большинство из клеток клона превращаются в плазматические клетки. Они перестают производить В-клеточный рецептор и начинают выделять полноценные растворимые антитела. Антитела попадают в кровь, разносятся по организму и связываются с антигеном. Где-то они просто его обезвреживают и в составе иммунных комплексов выводятся из организма. Где-то они работают опсонинами («черными метками») и активируют другие клетки иммунной системы.
После успешного иммунного ответа польза антител для организма не заканчивается. Часть клеток клона становится клетками памяти, они не участвуют в первичном иммунном ответе. Зато при повторной встрече с антигеном они помогут развить вторичный иммунный ответ. Им уже не понадобится искать «своего» Т-хелпера: при первой же встрече с антигеном они начинают производить антитела. Поэтому вторичный иммунный ответ развивается быстрее первичного и работает эффективнее. На этом основан эффект вакцинации — мы знакомим организм с новыми антигенами. Их можно вводить в составе убитого или ослабленного возбудителя заболевания или в виде отдельных молекул. Но принцип остается одним — мы вводим в организм антиген, он запускает иммунный ответ. Первичный ответ, как правило, будет слабым, так как антигена немного и он не вредит организму, но при этом формируется пул клеток памяти. И если через какое-то время приходится иметь дело с настоящим живым патогеном, то развивается вторичный ответ.
На измерении количества (титра) антител в крови основана диагностика многих заболеваний. Например, если мы хотим выяснить, есть ли в организме человека какая-нибудь бактерия, можно проверить кровь на антитела к ее ключевым антигенам. По количеству и типу антител мы можем сказать, насколько тяжело протекает заболевание и давно ли оно началось (в первичном ответе сначала участвуют IgM, потом IgG, во вторичном — в основном IgG, и их гораздо больше).
Какие еще бывают антитела
Можно представить себе антитело как конструктор: изменяя отдельные части молекулы, мы можем влиять на ее функцию. Изменится вариабельная часть — она станет специфична к другому антигену, изменится константная — ее станут узнавать другие иммунные клетки, и она сможет работать в других условиях (например, IgA — в слизистой оболочке). Этот принцип работает у всех позвоночных животных [11], и некоторые из них даже разработали полезные дополнения к этому конструктору, которые стоит взять на заметку [12].
У птиц репертуар антител менее разнообразен, чем у нас. В частности, у них нет отдельно взятых антител класса G и Е, зато есть промежуточный класс, сочетающий свойства и тех и других — IgY (Y — от yolk, «желток», поскольку этих антител много в яичном желтке). Молекулы IgY чуть тяжелее, чем наши IgG, и более жесткие. Зато их константную часть могут распознавать рецепторы и к IgG, и к IgЕ [13]. Если мы научимся создавать вакцины на основе куриных антител, то они будут лучше стимулировать иммунный ответ. И использовать их будет просто: берем курицу, заражаем ее каким-то патогеном (или просто его антигеном), а она откладывает нам яйца, богатые IgY, специфичным к этому патогену. Этот метод предполагают использовать для лечения разных воспалительных процессов, в том числе бактериальных инфекций. На данный момент он проходит клинические испытания, но конкретные препараты пока не разработаны.
Коровы пошли по пути удлинения антител. В их желудке обнаружены иммуноглобулины М со своеобразной «ручкой» — дополнительным отделом вариабельной части. Каждая «ручка» специфична к своему вирусному антигену. Полагают, что это уникальное изобретение жвачных для защиты пищеварительной системы. В их желудке пища задерживается надолго для переваривания желудочной микрофлорой. И для защиты этой микрофлоры от патогенов сформировались антитела с ручкой, которые могут дотянуться до эпитопов, спрятанных глубоко внутри молекулы антигена. Можно предположить, что, пришивая такую ручку к нашим иммуноглобулинам, мы сможем повысить их разнообразие, и они начнут связываться с эпитопами, до которых раньше не доставали. Но это пока теоретические размышления, до практических применений должно пройти еще немало времени.
А вот верблюды, сумчатые млекопитающие и хрящевые рыбы используют обратную стратегию — уменьшение антител (рис. 9). Среди их иммуноглобулинов есть фракция молекул, лишенных легких цепей, — их назвали HCAb (heavy chain antibody). Антигенсвязывающий участок у них образован только тяжелой цепью. Это легкое антитело оказалось очень выгодным, не зря оно появлялось в эволюции позвоночных несколько раз независимо. С одной стороны, оно легче, поэтому лучше путешествует по организму и выводится почками. С другой стороны, антигенсвязывающий участок меньше, поэтому может проникать, например, в активные центры ферментов. При этом антитело не только опсонизирует фермент, но и блокирует его работу. Это свойство ученые собираются использовать для борьбы против онкологических заболеваний. Можно от антитела верблюда отрезать константную часть, оставив только вариабельную — получится совсем маленькое нанотело. С помощью него можно, например, избирательно блокировать факторы роста раковых клеток [14], [15].
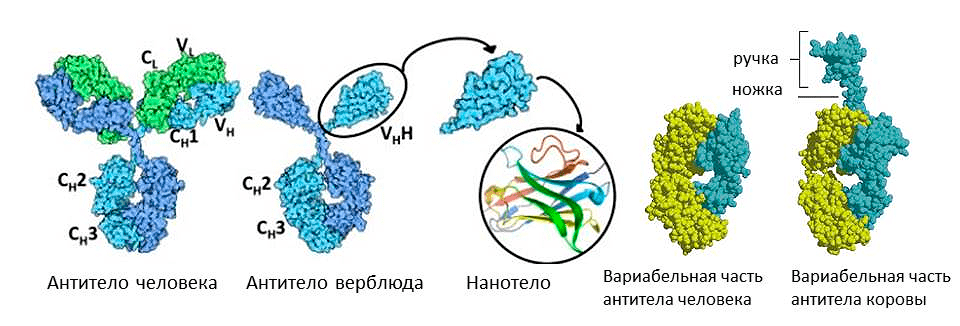
Рисунок 9. Необычные антитела у животных. Иммуноглобулины верблюда состоят из одних лишь тяжелых цепей (слева) а иммуноглобулин коровы несет дополнительную «ручку» на вариабельной цепи (справа).
Что мы можем сделать с антителами
Современные ученые не отстают от наших младших позвоночных братьев и тоже разрабатывают модифицированные антитела. Можно выделить два крупных направления работы с антителами. Первое — молекулярно-биологическое. Коль скоро антитела умеют специфично связываться с антигенами, можно использовать их как метку. Об этом мы подробно рассказывали в статье, посвященной иммунологическим методам [16]. Например, можно пришить к антителам светящиеся метки и окрашивать ими гистологические препараты: там, где мы увидим свечение, находится интересующее нас вещество.
Второе направление — медицинское [1]. С помощью антител можно не только иммунизировать человека, но и адресно доставлять в его организм вещества. Например, можно взять антитело, специфичное к белку опухоли, и пришить к нему (конъюгировать) токсичное вещество. Таким образом лекарство будет избирательно накапливаться в опухоли.
Антитела, безусловно, являются очень многообещающим конструктором. Однако для того, чтобы их широко применять, нужно научиться их производить в больших количествах. А для того чтобы ими лечить — убедиться в специфичности их действия и отсутствии побочных эффектов. О том, как лаборатории справляются с разработкой и производством антител, читайте в следующих статьях нашего цикла.
Департамент вычислительной биологии компании BIOCAD — спонсор спецпроекта
BIOCAD — международная биотехнологическая компания, которая создала умную технологическую платформу, объединившую в себе компьютерное моделирование и современные принципы синтеза генов de novo.

В компании реализован полный цикл выпуска лекарственных препаратов: от поиска лекарственной молекулы до массового производства и маркетинговой поддержки.
Компания ведет два масштабных проекта:
Для разработки лекарственных препаратов BIOCAD применяет технологию structure-based drug design, использующую методы компьютерного моделирования. Это позволяет сделать поиск молекул направленным. С помощью математического моделирования отобранная молекула оптимизируется под конкретную мишень, а затем воспроизводится в реальной лаборатории.
В основе большинства разработок компании лежит математическое моделирование. То, что раньше было возможно осуществить исключительно in vitro в стенах лабораторий, по мнению исследователей BIOCAD, сегодня может быть воплощено in silico силой чистого разума.
Здесь собрали одну из лучших команд биоинформатиков в стране, которая занимается научными исследованиями, разрабатывает и внедряет новейшие методы интеллектуального анализа данных. В ее распоряжении один из мощнейших вычислительных кластеров, и если еще 2–3 года назад можно было только мечтать о решении задач направленного дизайна белковых молекул, то сейчас это одно из направлений работ Департамента вычислительной биологии.
Материал предоставлен партнёром — Департаментом вычислительной биологии компании BIOCAD