Не только ВИЧ. Как устроены вирусы?
Галина Вирясова
Сайт СПИД.ЦЕНТР немало рассказывает своим читателям о вирусе иммунодефицита человека, но как устроены остальные вирусы? Чтобы помочь увидеть общую картину, наши авторы подготовили небольшой гид по вирусам с объяснением, как они работают, размножаются и как работают противовирусные лекарства.
Одни вирусы способны интегрироваться в геном клетки-мишени и таким образом оставаться во всех дочерних клетках, которые будут в будущем получены после ее деления. К таким вирусам относятся гаммаретровирусы и лентивирусы. Другие делать этого не умеют (например, адено- и аденоассоциированные вирусы). Но для производства белков и репликации (размножения) все они используют клетку и ее синтетический аппарат.
Несмотря на некоторую «несамостоятельность» в размножении, вирусы способны наследовать генетические мутации и подвержены эволюционному отбору. Выживает сильнейший, а в случае вируса — самый устойчивый и заразный.
Как это работает?
Для того чтобы вирус мог проникнуть в клетку, белки его оболочки должны связаться с мембранными белками клетки-мишени. Важно отметить, что проникает вирус только в те клетки, которые могут в дальнейшем помочь его репликации. Вирус ВИЧ живет в клетках иммунной системы, вирус гепатита С — в клетках печени. Есть особые вирусы, которые поражают только растения или даже только бактерии.
В целом у вирусов существуют разные стратегии доставки вирусного материала в клетку. Какие-то вирусы размножаются в ее цитоплазме, а какие-то — в ядре. Некоторые умеют «впрыскивать» свою генетическую информацию прямо через мембрану, когда сам капсид остается снаружи.
Объединяет их одно: после того как вирусная информация доставлена в клетку, та, «забывая» о своей изначальной функции, начинает заниматься в первую очередь репликацией вируса. Клетка производит матричную РНК (мРНК), с которой затем синтезируются вирусные белки и копируется геном, и сама собирает новую вирусную частицу.
В большинстве случаев вирус убивает клетку, чтобы выйти наружу и приступить к поиску новой «жертвы». Но иногда этого не происходит: некоторые вирусы, в том числе ВИЧ, могут отделяться от клетки, обзаведясь собственной оболочкой и оставив клетку в живых, чтобы та продолжила производить новые вирусные частицы.
Содержащие неактивный вирус и оставшиеся в живых клетки иногда сохраняют возможность нормального функционирования. В этом случае клетки могут быть заражены, но вирус проявит себя спустя длительный период времени. Так устроен герпес.
В зависимости от того, каким типом нуклеиновой кислоты представлен генетический материал, выделяют ДНК-содержащие вирусы и РНК-содержащие вирусы. И тут стоит остановиться на классификации.
Типы вирусов. Коротко о главном
Современная типология вирусов содержит 7 классов и была предложена Дэвидом Балтимором еще в 1971 году. С тех пор, впрочем, она была уточнена и расширена, в том числе советскими учеными. И выглядит в настоящее время таким образом:
Вирусы, содержащие двухцепочечную ДНК
Описание
Для репликации вирусу необходимо попасть в ядро клетки-мишени и воспользоваться ее ДНК-полимеразой. Иногда вирус вызывает незапланированное деление самое клетки, то есть становится онкогенным. Эти вирусы хорошо изучены.
Пример: Вирус герпеса, адено- и папилломавирусы
Вирусы, содержащие одноцепочечную ДНК
Описание
Попадая в ядро клетки, вирусы образуют двухцепочечную ДНК, после чего реплицируются так же, как вирусы класса I.
Пример: Парво- и цирковирусы
Вирусы, в которых РНК способна к репликации (редупликации)
Описание
Вирусы этого класса могут размножаться в цитоплазме клетки, им не нужна молекула ДНК. Каждый ген, находящийся в РНК вируса, кодирует только один вирусный белок.
Пример: Бирна- и реовирусы
Вирусы, содержащие одноцепочечную (+) РНК
Описание
Из геномной (+) РНК на рибосомах хозяина создаются вирусные белки. В одном фрагменте РНК могут быть закодированы разные белки, что увеличивает сложность вируса без удлинения генов.
Пример: Пикорнавирусы (полиомиелит, гепатит А) и коронавирусы
Вирусы, содержащие одноцепочечную (–) РНК
Описание
(–) РНК этих вирусов предварительно должна быть транскрибирована в (+) РНК вирусными РНК-полимеразами, после чего может начаться синтез вирусных белков. Вирусы этого класса делятся еще на две группы, в зависимости от их генома и места его репликации (цитоплазма или ядро).
Пример: Филовирусы, аренавирусы (геморрагическая лихорадка Ласса), ортомиксовирусы (вирусы гриппа) и так далее.
Вирусы, содержащие одноцепочечную (+) РНК, реплицирующиеся через стадию ДНК
Описание
Такие вирусы используют фермент обратную транскриптазу для превращения (+) РНК в ДНК, которая встраивается в геном хозяина ферментом интегразой. Дальнейшая репликация происходит при помощи полимераз клетки хозяина.
Пример: Ретровирусы (в том числе ВИЧ)
Вирусы, содержащие двухцепочечную ДНК, реплицирующиеся через стадию одноцепочечной РНК
Описание
Молекула ДНК замкнута в кольцо и является матрицей для синтеза мРНК и дополнительных молекул РНК, которые используются при репликации вирусного генома обратными транскриптазами.
Пример: Колимовирусы (вызывают инфекции растений) и гепаднавирусы (например, гепатит В)
Вакцинация и лечение
Как правило, организмы умеют бороться с паразитирующими на них вирусами. На примере млекопитающих и человека мы обычно говорим о главном инструменте — врожденном иммунитете.
Впрочем, наиболее эффективен этот вид защиты в отношении бактериальных инфекций и не может обеспечить продолжительную и надежную защиту, особенно от инфекций вирусных.
Именно поэтому огромное значение имеет приобретенный иммунитет, в результате которого клетки иммунной системы обучаются вырабатывать специфические к вирусу антитела, способные уничтожать как саму вирусную частицу, так и зараженные ею клетки.
Еще одна врожденная система борьбы с вирусными инфекциями — внутриклеточная. Как правило, клетка способна распознать чужеродную РНК в своей цитоплазме, куда ее сперва и доставляют многие вирусы, и имеет специальные комплексы для ее деградации. Но часть вирусов научились обходить и эту ловушку. К примеру, ротавирусы, которые даже внутри клетки сохраняют капсид с геномной РНК.
С приобретенным иммунитетом тоже не все гладко. Некоторым вирусам, например, ВИЧ, удается избежать иммунного ответа. Другим, например нейротропным вирусам, — уклониться от него, выбрав безопасную среду обитания: они распространяются среди клеток нервной системы, где их не может «достать» иммунная система. Самый известный из таких вирусов — вирус бешенства, который способен проникать в нейроны.
Миссия: уничтожить
Основная сложность в лечении вирусных заболеваний заключается в том, что они используют естественные функции клеток-мишеней для своего размножения, поэтому ученым зачастую оказывается не так-то просто придумать препарат, который будет токсичен для вируса и безопасен для самой клетки. Если такой безопасности достичь не удастся, лекарство будет иметь слишком много побочных эффектов, повреждающих сам организм, что окажется нецелесообразно для использования.
По принципу действия противовирусные препараты подразделяются на две группы: стимулирующие иммунную систему атаковать вирусы (например, за счет индукции синтеза белков-интерферонов) и атакующие вирусы напрямую. Препараты второй группы различаются по этапу жизненного цикла вируса, на котором они активны: это препараты, препятствующие проникновению вируса в клетку, препятствующие размножению вируса внутри клетки и препятствующие выходу копий вируса из клетки.
Чтобы помешать проникновению вируса, препарат должен заблокировать рецептор на клетке, с которым связывается вирусная частица. Так работает, например, ибализумаб — зарегистрированный в США новый препарат против ВИЧ, о котором мы недавно писали.
Такие противовирусные препараты, как уже давно известный ацикловир (им лечат инфекции, вызванные простым вирусом герпеса) или ламивудин (активен против ВИЧ и гепатита В), представляют собой синтетические аналоги нуклеозидов — «букв», из которых состоят нуклеиновые кислоты. Если эти модифицированные, неправильные нуклеозиды попадают в клетку, вирусный геном, в который они оказались встроены, становится непригоден для дальнейшего распространения вируса.
Еще один класс противовирусных препаратов блокирует ферменты, необходимые для создания и модификаций белков вируса. Такие лекарства называют протеазными ингибиторами.
Вместо заключения: а могут ли вирусы приносить пользу?
Безусловно, да. Несмотря на то, что вирусы ассоциируются у большинства людей с однозначным вредом, они могут приносить и пользу — если речь идет о так называемых вирусных векторах и терапевтических подходах на их основе.
Исследователи давно научились помещать в белковую оболочку вируса интересующие их нуклеиновые кислоты, чтобы доставлять нужный ген в клетки, а также убирать те гены, которые делают вирус опасным для организма.
Это позволило сделать возможной генную терапию, помогающую бороться с заболеваниями, вызванными известными генетическими мутациями. Создание вирусных векторов — достаточно непростая задача, к тому же ограниченная свойствами самих вирусных частиц: количеством помещающейся генетической информации, местом ее вставки, стабильностью. Кроме того, вирусный вектор, используемый в медицине, не должен вызывать иммунного ответа или критично влиять на жизнедеятельность клетки. Тем не менее эти сложности решаются, поэтому уже одобрен ряд вполне успешных и безопасных генных терапий. А в качестве основы для вирусных векторов чаще всего используются ретро-, ленти-, адено- и аденоассоциированные вирусы.
Вирусные геномы в системе эволюции
Человеческая Т-клетка (синий), атакованная ВИЧ (желтый). Вирус ориентирован на Т-клетки, которые играют важную роль в иммунной реакции организма против вторжений, таких как бактерии и вирусы.
Автор
Редакторы
Статья на конкурс «био/мол/текст»: Изучение виромов (популяций вирусов, объединенных общей средой обитания) практикуется в современной вирусологии всё чаще. Подобный подход приносит неожиданные результаты — концентрация вирусов в природе, оказывается, чрезвычайно высока. Например, в морской среде количество вирусных частиц превосходит количество клеток в разы. При этом вирусные частицы способны к передаче генетической информации. Исследования, опубликованные в 2010 году, показали, что агенты переноса генов (АПГ, особая разновидность вирусоподобных частиц, участвующих в горизонтальном переносе генов) привносят в геном бактерий новые фрагменты значительно чаще, чем считалось ранее. А если вирусные частицы настолько распространены в биосфере, и большая часть из них способна к переносу генов, то важно рассмотреть вирусы в рамках общей модели эволюции.

Конкурс «био/мол/текст»-2014
Эта статья представлена на конкурс научно-популярных работ «био/мол/текст»-2014 в номинации «Биоинформатика и молекулярная эволюция».

Главный спонсор конкурса — дальновидная компания «Генотек».
Конкурс поддержан ОАО «РВК».
Спонсором номинации «Биоинформатика» является Институт биоинформатики.
Спонсором приза зрительских симпатий выступила фирма Helicon.
Свой приз также вручает Фонд поддержки передовых биотехнологий.
Вопрос о происхождении вирусов
Существует три основные теории возникновения вирусов:
Однако фундаментальные достижения, такие как открытие рибозимов (молекул РНК, обладающих каталитическим действием), разработка гипотезы о «мире РНК» [2], открытие новых форм вирусов и успехи в геномике делают наименее популярную в прошлом гипотезу доклеточного происхождения все более весомой. И то, что некоторые вирусные белки не обнаруживают гомологии с белками бактерий, архей и эукариот [3], свидетельствует о сравнительно давнем обособлении этой группы. В остальном же достоверно объяснить происхождение вирусов на основании трёх закрепившихся классических гипотез пока еще не вполне удаётся.
Зарождение жизни. Идея последнего универсального общего предка: каким он мог бы быть и что ему предшествовало?
Как известно, концепцию трехдоменной классификации организмов предложил Карл Вёзе [4] в 1977 году, разделив современный биологический мир на три домена — бактерии, археи и эукариоты. Обычно ее представляют в виде дерева с тремя «ветвями», имеющими общий «ствол» (рис. 1). Подобные взгляды были близки еще Дарвину, который видел в подобной концепции логичное завершение своей теории. Так появился термин «Последний универсальный общий предок» (англ. last universal common ancestor, LUCA) — ближайший общий предок всех ныне живущих на Земле живых организмов.
Некоторые ученые считают, что можно выделить и четвертый домен жизни: «Гигантские вирусы: 4-й домен жизни?» [5]. — Ред.
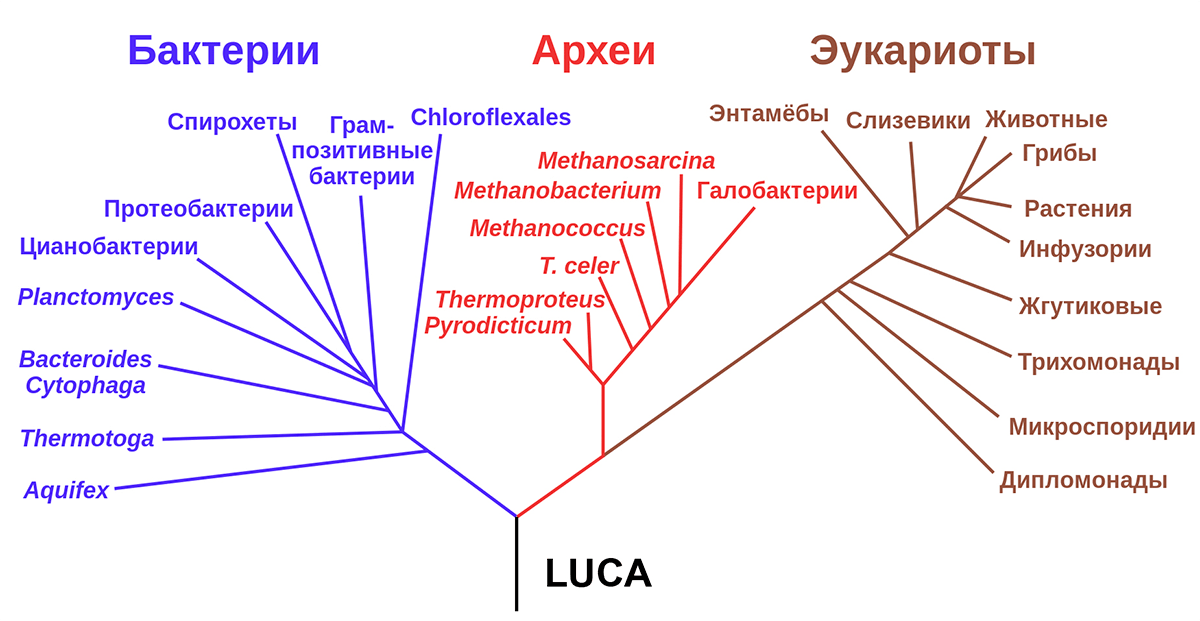
Рисунок 1. Схема трехдоменной классификации, предложенная Вёзе. В основании этой схемы должен находиться последний универсальный общий предок (англ. last universal common ancestor, LUCA).
Самый сильный аргумент в пользу существования LUCA — сохранившаяся общая система экспрессии генов (передачи наследственной информации от гена с образованием РНК или белков), одинаковая для всех живущих организмов. Все известные клеточные формы жизни используют один и тот же генетический код из 20 универсальных аминокислот и стоп-сигналов, закодированных в 64 кодонах (единицах генетического кода). Трансляция генетической информации в процессе синтеза белков по заданной матрице выполняется рибосомами, состоящими из трех универсальных молекул РНК и примерно 50 белков, из которых 20 так же одинаковы для всех организмов.
В 2010 году американский биохимик Даглас Теобальд математически проверил вероятность существования LUCA [6]. Он выбрал 23 белка, встречающихся у организмов из всех трех доменов, но имеющих разную структуру у различных видов. И исследовал эти белки у 12 различных видов (по четыре из каждого домена), после чего использовал компьютерное моделирование различных эволюционных сценариев, чтобы понять, при каком из них наблюдаемая картина будет наиболее вероятной. Оказалось, что концепция, включающая существование универсального предка, значительно вероятнее концепций, где его нет. Еще более вероятна модель, основанная на существовании общего предка, но допускающая обмен генами между видами [7].
Предположение о том, что LUCA был прокариотической клеткой, похожей на современные, часто принимается по умолчанию. Однако мембраны архей и бактерий имеют разное строение (рис. 2). Получается, что общий предок должен был обладать комбинаторной мембраной. Новая информация о мембранах LUCA появилась в 2012 году, когда несколько групп ученых подробно проанализировали историю генов всех ферментов биосинтеза компонентов липидов у бактерий, архей и эукариот [8].
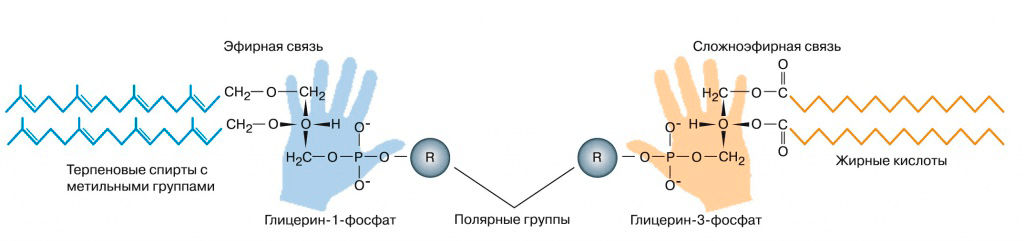
Рисунок 2. Строение мембранных липидов бактерий (справа) и архей (слева)
Родственными у архей и бактерий оказались ферменты для синтеза терпеновых спиртов и пришивания полярных голов к спиртам. Значит, эти реакции мог проводить и LUCA. Проще всего было предположить, что липиды LUCA состояли из одного остатка терпенового спирта, остатка фосфата и полярной группы (серина или инозитола). Подобные липиды были синтезированы искусственно. Образующиеся из них мембраны обладают высокой подвижностью по сравнению с современными мембранами, хорошо пропускают ионы металлов и малые органические молекулы. Это могло позволять древним протоклеткам поглощать готовую органику из внешней среды даже без транспортных белков.
Реконструкции LUCA методами сравнительной геномики указывают на то, что это должен быть сложный организм без обширного ДНК-генома (геном, состоящий из нескольких сотен РНК-сегментов или ДНК провирусного типа). Но даже если считать возможность существования общего предка доказанной, остается загадкой, в какой среде он мог бы появиться.
«Относительная генетическая сложность могла быть свойственна клеткам еще до появления современного типа клеточной организации — для этого сценария мы будем использовать обозначение LUCAS (рис. 3) (Last Ancestral Universal Common State — последнее универсальное предковое состояние [всего живого]». — пишет биолог Евгений Кунин в своей книге «Логика случая. О природе и происхождении биологической эволюции» [9].

Рисунок 3. Сценарий вирусного мира в гипотезе доклеточного происхождения вирусов подпись
Предполагается, что идеальные условия для формирования жизни существовали вблизи термальных геоисточников (морских или наземных) в виде сети неорганических ячеек, обеспечивающих градиенты температуры и рН, способствующих первичным реакциям, и предоставляющих универсальные каталитические поверхности для примитивной биохимии [10].
Эти отсеки могли быть населены разнородной популяцией генетических элементов. Вначале сегментами РНК. Затем более крупными и сложными молекулами РНК (один или несколько белок-кодирующих генов). А позднее и сегментами ДНК, которые постепенно увеличивались (рис. 3).
Такие простейшие генетические системы использовали неорганические соединения из раствора и продукты деятельности других генетических систем. Сначала они должны были подчиняться индивидуальному отбору ввиду большого разнообразия. Но ясно, что важным фактором такого отбора была способность передавать генетическую информацию, то есть, копировать себя. Присутствие одновременно в одной ячейке молекул, способных копировать РНК, кодировать полезные белки и управлять синтезом новых молекул, давало больше шансов выживать в каждой отдельной ячейке. И в такой системе рано или поздно должны были появиться паразитирующие элементы. А если это так, то вирусные элементы стоят у самых истоков эволюции [11].
Возникновение паразитов — неизбежное последствие эволюционного процесса
Если рассматривать эволюцию как единый процесс, становится понятно, что виды никогда не эволюционируют независимо друг от друга. Более того, паразиты появляются и в компьютерных моделях эволюции простых репликаторов (самовоспроизводящихся РНК-подобных систем) [9]. Так исследования, проведенные в 2008 году Набуто Такеучи и Паулин Хогвег, представляют собой расчет такой репликативной системы [12]. Простота системы позволяет явно смоделировать отображение генотип-фенотип взаимодействий отдельных репликаторов. Результаты показали, что популяция репликаторов, первоначально состоящая из одного генотипа, развивается в сложную экосистему до четырех «видов». В этой диверсификации «виды» эволюционируют путем приобретения уникальных генотипов с отличной экологической функциональностью, причем два из четырех «видов» становятся паразитами, неспособными к самостоятельной репликации.
Анализ диверсификации показывает, что паразитарные репликаторы, которые были придуманы, чтобы дестабилизировать разнообразие репликатора, на самом деле способствуют эволюции разнообразия посредством создания новой «ниши» для каталитических репликаторов (самовоспроизводящихся репликаторов). Это также делает существующую систему репликатора чрезвычайно стабильной при эволюции паразитов, поскольку в такой системе паразиты имеют преимущество репликации на катализаторах.
Дело в том, что репликация катализаторов требует некоторого периода времени, и паразиты, которые не копируют другие молекулы, могут тратить больше времени на репликацию, чем катализаторы. Таким образом, присутствие катализаторов влечет за собой «нишу» для паразитов. В нынешней системе (рис. 4), C-катализатор (cat-C) создает такую нишу, и это позволяет эволюционировать G-паразиту (par-G). Кроме того, как только создается структура C-катализатор-G-паразит, она создает еще один нишу для фенотипа, который может высвободиться из G-паразита. И если такой фенотип приобретает А-катализатор (cat-A), он тоже эволюционирует. Наконец, создание такого альтернативного катализатора создает нишу для фенотипа, который мог бы паразитировать на этом альтернативном катализаторе (par-U).
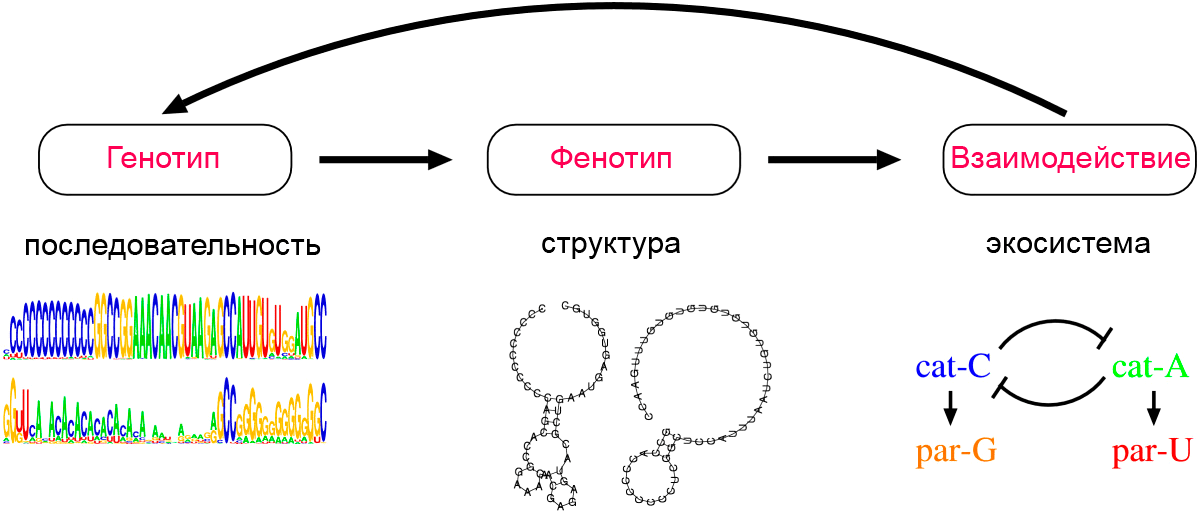
Рисунок 4. Схематическое представление структуры модели эволюции РНК-подобной системы. На втором этапе цепочки последовательностей начинают соединяться комплементарными связями сами с собой. В результате у двух видов (cat-C и cat-A) возникает вторичная структура молекулы, которая обладает каталитическим свойством. Она ускоряет собственную репликацию (или репликацию несвернувшихся соседей). Два вида при этом приобретают паразитические свойства (par-G и par-U). Пояснения в тексте.
Таким образом, паразитарные репликаторы способствуют эволюции разнообразия, вместо того, чтобы мешать этому разнообразию. Это также делает существующую систему репликатора чрезвычайно стабильной при эволюции паразитов.
Подобную «гонку вооружений» можно наблюдать, если на молекулярном уровне изучить системы «ионный канал — нейротоксин», которые коэволюционируют, видимо, как раз по этому принципу: «Яды — высокоточное оружие: компьютерное исследование природных нейротоксинов» [13]. — Ред.
Механизмы клеточной защиты против вирусов
Прежде чем говорить о механизмах взлома и защиты подробнее, следует отметить, что вирусы в большинстве случаев не имеют цели убивать клетки хозяев, в которые они попали. У них даже нет цели навредить — это невыгодно для самого вируса. Однако если дело обстоит именно так, какова же причина болезней и эпидемий, вызванных вирусами? Ведь часто при заражении вирусной инфекцией клетка-носитель погибает или становится недееспособной. Конечно, попадая в клетку, вирус перенаправляет ресурсы клетки на свои нужды. Многие вирусы запрещают экспрессию генов клетки-хозяина, что играет главную роль в способности вируса вызывать болезнь. Но наибольший вред исходит от нерасчетливых защитных действий хозяев. А также от противозащитной активности вирусов, не связанных напрямую с механизмами вирусной репродукции. Другими словами, основной вред оказывает не размножение вируса как таковое, а противоборство клеточной защиты и постоянно меняющейся вирусной противозащиты. Само присутствие вируса не всегда сказывается на функциональной работе клетки [14], [15]. Например, в фитопаталогии (науке о болезнях растений) существует термин «толерантность» — патогенный вирус может активно размножаться в растении, не вызывая болезненных проявлений.
Механизмы защиты от вирусов стандартны, поскольку все вирусы уникальны, и приспособиться к каждому не представляется возможным. Это такие механизмы как:
Получается, что клетка борется с вирусом, нарушая собственные обмен веществ и/или структуру. Защитные реакции клетки — это в основном самоповреждающие механизмы.
Вирус заражает конкретную клетку потому, что его механизмы нападения направлены именно против данного типа клеток. Это такие механизмы как:
Схемы защитных приемов клетки и противозащиты вирусов во многом идентичны. Вирусы и клетки применяют одни и те же приемы. Для подавления синтеза вирусных белков клетка использует интерферон, а чтобы подавить образование интерферона, вирус угнетает синтез белков.
Поскольку узнавание вируса неспецифическое, клетка не может знать намерения конкретного вируса. Она может бороться с вирусом лишь стандартными приемами, поэтому ее оборонные действия часто могут быть чрезмерными.
«Можно сказать, что человек, умерший от полиомиелита, погибает от того, что его организм слишком сильно боролся с вирусом. Будь он аккуратнее в этой борьбе, вероятно, выжил бы», — вот слова прославленного русского вирусолога В.И. Агола.
Понятие о вирусном геноме, типы вирусных генов, концепция генов-сигнатур
В исследовании, проведенном вирусологом Евгением Куниным и его коллегами [16], анализ последовательностей вирусных геномов выявил несколько категорий вирусных генов, принципиально отличающихся по происхождению. Можно обсуждать, какая степень дробности классификации оптимальна, но четко различаются пять классов, укладывающихся в две более крупные категории.
Гены с четко опознаваемыми гомологами у клеточных форм жизни:
Вирусные сигнатуры — это гены, общие для многих разнообразных групп вирусов, имеющие лишь отдаленные гомологи у клеточных организмов, для которых имеются убедительные свидетельства в пользу монофилии (общего происхождения) всех вирусных членов соответствующих семейств генов [9]. Фактически эти гены являются признаками «вирусного состояния». Гены-сигнатуры предшествуют клеткам и происходят непосредственно из первичного, доклеточного пула генов.
Таким образом, отличительные особенности генов-сигнатур:
Из всего вышесказанного следует, что эти гены переходили от вируса к вирусу (или к элементу, подобному вирусу) на протяжении четырех миллиардов лет эволюции жизни, а вирусные геномы появились благодаря перемешиванию и подгонке друг к другу генов в гигантской генетической сети, которую представляет собой мир вирусов. Многочисленные гены клеточных форм жизни также пронизывают эту сеть, прежде всего благодаря геномам крупных вирусов, таких как NCDLV и крупным бактериофагам, которые позаимствовали множество генов от своих хозяев на разных этапах эволюции. Однако большинство заимствованных генов сами по себе не критичны для репликации и экспрессии вирусного генома (исключая некоторые случаи возможного неортологичного замещения генов-сигнатур); обычно эти гены участвуют во взаимодействии между вирусом и хозяином. Таким образом, несмотря на интенсивный взаимообмен генами с хозяевами, вирусы всегда происходят от других вирусов.
Вирусы, встроенные в геном, и горизонтальный перенос генов
В процессе эволюции многие вирусы встроились в геномы клеточных форм жизни путем горизонтального переноса генов (ГПГ). Впервые горизонтальный перенос был описан в 1959 году, когда ученые продемонстрировали передачу резистентности к антибиотикам между разными видами бактерий. В 1999 году Рави Джайн, Мария Ривера и Джеймс Лейк в своей статье писали о произошедшей значительной передаче генов между прокариотами [17]. Этот процесс, по-видимому, оказал некоторое влияние также и на одноклеточные эукариоты. В 2004 году Карл Вёзе опубликовал статью, в которой утверждал, что между древними группами живых организмов происходил массивный перенос генетической информации. В древнейшие времена преобладал процесс, который он называет горизонтальным переносом генов. Причем, чем дальше в прошлое, тем это преобладание сильнее [18].
Современные же данные показывают, что ГПГ имеет огромное влияние не только на эволюцию простейших, но и на эволюцию многоклеточных. Молекулярный биолог Петер Гогартен описал горизонтальную генную передачу как «новую парадигму биологии».
Горизонтальный перенос генов — процесс, в котором организм передаёт генетический материал другому организму, не являющемуся его потомком. Горизонтальная передача генов реализуется через различные каналы генетической коммуникации — процессы конъюгации, трансдукции, трансформации, переноса генов в составе плазмидных векторов, вирусов, мобильных генетических элементов (МГЭ).
Трансдукция — перенос бактериофагом (агентами переноса генов, АПГ) в заражаемую клетку фрагментов генетического материала клетки, исходно содержавшей бактериофаг [19]. Такой бактериофаг обычно переносит лишь небольшой фрагмент ДНК хозяина от одной клетки (донор) к другой (реципиент). В зависимости от типа трансдукции — неспецифической (общей), специфической или абортивной, геном фага или хозяина-бактерии может быть изменен тем или иным образом:
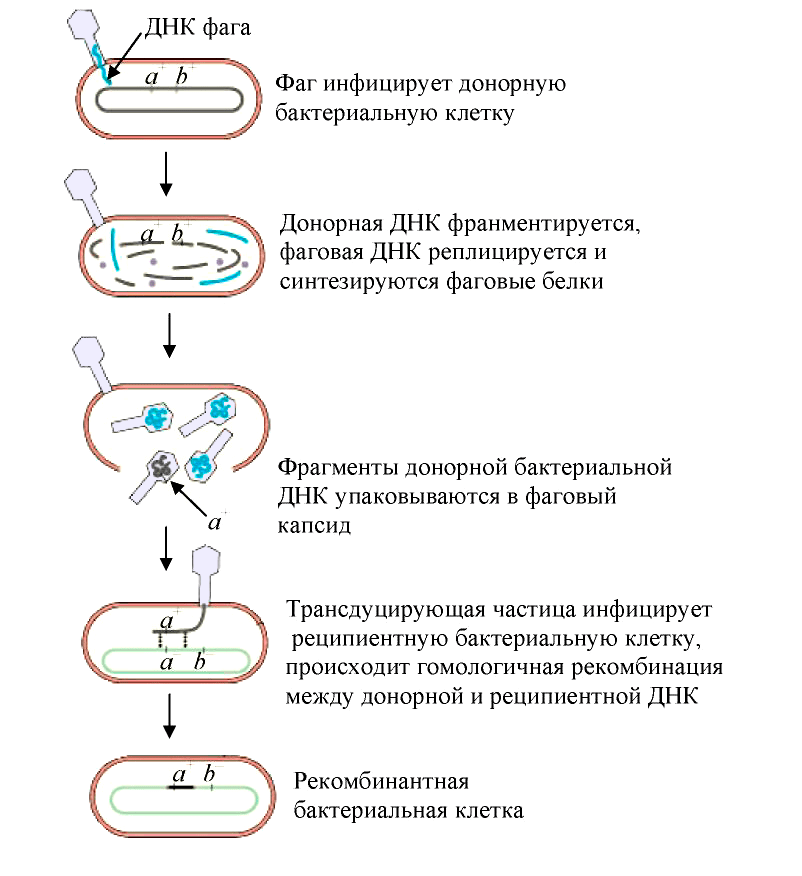
Рисунок 5. Схема общей трансдукции
Наиболее известным примером специфической трансдукции служит трансдукция, осуществляемая фагом λ. Поскольку этот фаг при переходе в состояние профага включается в хромосому бактерий между генами, кодирующими синтез галактозы и биотина, именно эти гены он может переносить при трансдукции.
Важнейшую роль в эволюции животных играют гены, которые животные заимствуют у вирусов, транспозонов, ретротранспозонов, дельта-ретровирусов (экозогенные и эндогенные вирусы). Фрагменты ДНК вирусов и транспозонов часто «приручаются» высшими организмами и начинают выполнять полезные функции в геноме. Большинство встроенных (ретровирусных, если говорить о высших позвоночных) геномов совершенно бесполезны для хозяина. Фактически идет постоянный процесс превращения так называемой «мусорной» или «эгоистической» ДНК вирусного происхождения в полезные элементы генома. Явление это настолько широко распространено, что для него даже предложен специальный термин — молекулярное одомашнивание [21].
Насколько действительно мусорна мусорная ДНК, обсуждается в статье «Сколько сора в нашей ДНК» [20]. — Ред.
Вот несколько примеров важных эволюционных событий, связанных с молекулярным одомашниванием:
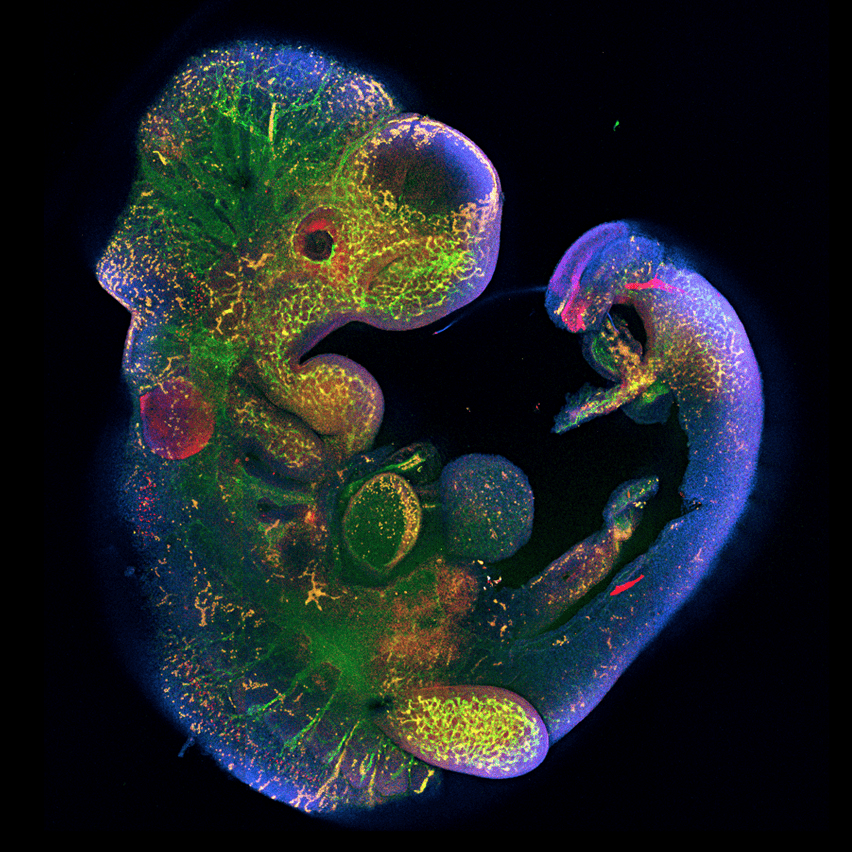
Рисунок 6. Роль гена Peg10 в эмбриональном развитии. Ученые под руководством Рюичи Оно из Токийского медицинского университета Японии показали, что у мышей с выключенным геном Peg10 нарушается развитие плаценты, от чего эмбрион погибает через 10 дней после зачатия [24].
В 2008 году в ходе целенаправленного поиска неиспорченных вирусных генов в геноме человека исследователи нашли два очень похожих друг на друга ретровирусных гена (их назвали ENVV1 и ENVV2), которые, по всей видимости, находятся в рабочем состоянии [25]. Это гены белков оболочки ретровируса. Каждый из них входит в состав своего эндогенного ретровируса (ЭРВ), причем все остальные части этих ЭРВ давно не функционируют.
Авторы нашли гены ENVV1 и ENVV2 и в геномах других приматов: у человекообразных (шимпанзе, орангутана) и других обезьян Старого Света (макаки-резуса, зеленой мартышки), а также у обезьян Нового Света — мармозетки и беличьей обезьяны. Однако у наших более отдаленных родственников (лемуры) этих генов нет. Сопоставив данные по нуклеотидным последовательностям генов ENVV1 и ENVV2 у разных обезьян, исследователи смогли реконструировать эволюционную историю этих генов. В эволюции приматов имело место приобретение полезных генов «со стороны», а именно от ретровирусов. По своему происхождению они являются генами белков оболочки ретровируса [21]. Исходный ретровирус встроился в геном наших предков и стал эндогенным ретровирусом после того, как разделились линии обезьян и лемуров, но до того, как разошлись обезьяны Старого и Нового света, то есть примерно 43–77 млн. лет назад. Этот ЭРВ вскоре подвергся двум последовательным дупликациям, и в результате получилось три одинаковых ЭРВ, расположенных по соседству на одной хромосоме. Все участки этих ЭРВ, кроме генов белков оболочки, стали постепенно дегенерировать. Ген ENVV2 приобрел в результате мутаций какое-то полезное для хозяина свойство и стал сохраняться отбором. Другие два гена, по-видимому, тоже приобрели полезные функции, но они оказались не столь незаменимыми, как ENVV2. Поэтому в некоторых эволюционных линиях обезьян эти гены были утрачены или выведены мутациями из строя. В частности, общий предок человека и шимпанзе потерял ген ENVV3.
Вирусные гены ENVV1 и ENVV2 у человека и обезьян работают в плаценте и, скорее всего, выполняют следующие функции:
Таким образом, как минимум три полезных применения нашли себе вирусные гены в плаценте приматов. Это показывает, что генетические модификации, которым ретровирусы подвергают организмы, в долгосрочной перспективе могут оказаться полезными или даже определить развитие вида. И с учетом всего вышесказанного древо доменов должно выглядеть как на схеме ниже (рис. 7).
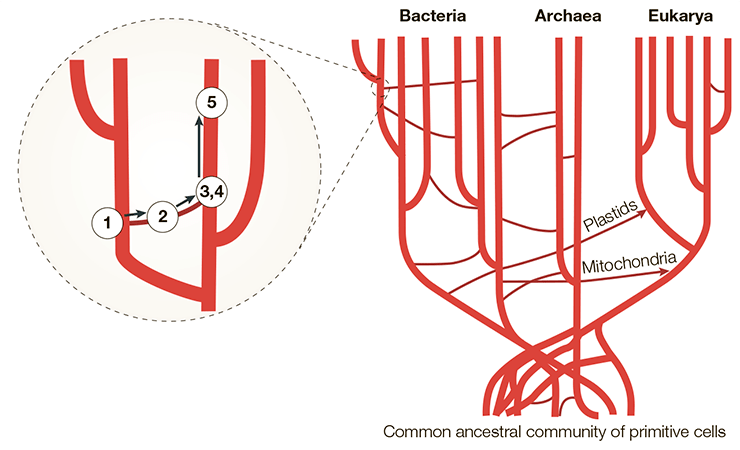
Рисунок 7. Горизонтальный перенос генов в рамках трехдоменного дерева
Заключение
Возникновение паразитов — обязательная черта эволюционирующих систем репликаторов, а соревнование хозяев и паразитов движет эволюцию тех и других. Любой организм является результатом миллионов лет борьбы клеток с невероятно разнообразным миром вирусов. Их действия и их эволюция пронизывают всю историю клеточной эволюции, и сейчас меняется само наше представление о них. Когда-то вирусы считали деградировавшими клетками, но чем больше мы узнаем о вирусах, тем очевиднее, что их роль в общей эволюции значительна. И невероятно много нам еще предстоит узнать.
Статья написана в соавторстве с Евгенией Щепенок.