Функции ретикулярной формации
Ретикулярная формация не осуществляет специфические, какие-то определенные рефлексы Функция РФ иная.
1. Во-первых, РФ обеспечивает, интеграцию, объединение функций всей ЦНС. Она является главной интегрирующей, ассоциативной системой ЦНС. Эту функцию она выполняет потому, что РФ, ее нейроны образуют огромное количество синапсов как между собой, так и с другими отделами ЦНС. Поэтому, возбуждение попав в РФ, очень широко распространяется, иррадиирует и по ее эфферентным путям: восходящим и нисходящим это возбуждение достигает всех частей ЦНС. В результате этой иррадиации включаются и вовлекаются в работу все образования ЦНС, достигается содружественная работа отделов ЦНС т.е. РФ обеспечивает формирование целостных рефлекторных реакций, в рефлекторной реакции участвует вся ЦНС
II. Вторая функция РФ заключается в том, что она поддерживает тонус ЦНС, т.к. РФ сама всегда находится в тонусе, тонизирована. Её тонус обусловлен рядом причин.
1).РФ обладает очень высокой хемотропностью Здесь находятся нейроны обладающие высокой чувствительностью к некоторым веществам крови (например, к адреналину, СО;) и лекарствам (к барбитуратам, аминазину и др.).
3). Тонус РФ поддерживается также за счет импульсов, которые идут по нисходящим путям от коры больших полушарий, от базальных ганглиев.
4). В поддержании тонуса сетчатого образования имеет большое значение также длительная циркуляция нервных импульсов в самой РФ, имеет значение реверберация импульсов в РФ. Дело в том, что в РФ находится громадное количество нейронных колец и по ним информация, импульсы циркулируют часами.
5). У нейронов РФ длительный латентный период ответа на периферическую стимуляцию в связи с проведением возбуждения к ним через многочисленные синапсы.
6). Они имеют тоническую активность, в покое 5—10 имп/с.
В результате выше названных причин РФ всегда находится в тонусе и от нее импульсы поступают в другие отделы ЦНС. Если перерезать ретикуло-кортикальные пути, т.е. восходящие пути, идущие от РФ в кору, то кора головного мозга выходит из строя, так как она лишилась главного источника импульсов.
3.2.2. Ретикулярная формация продолговатого мозга
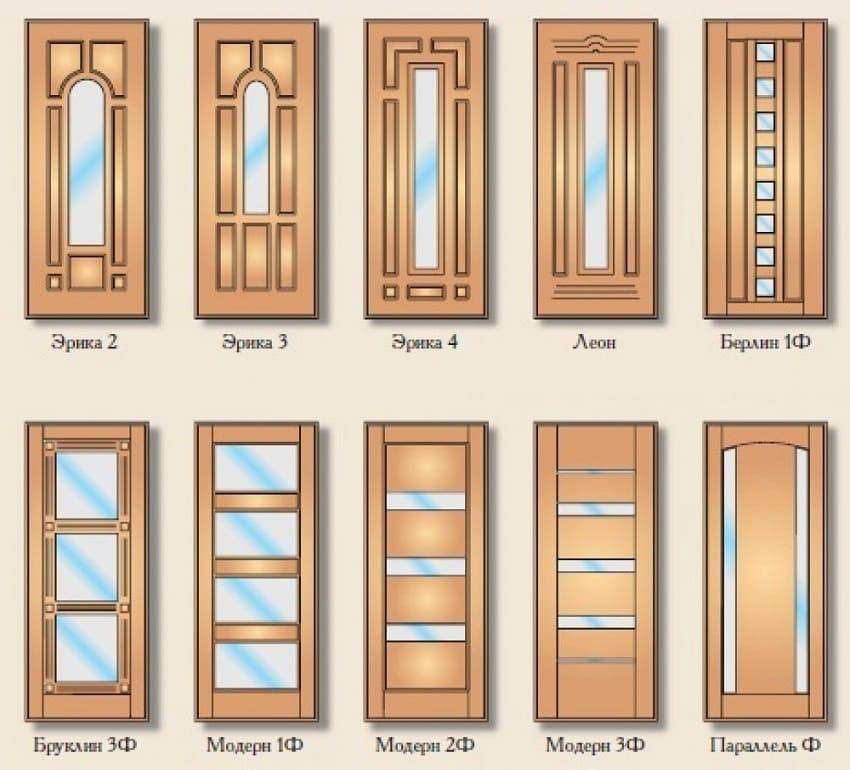
Ретикулярная формация образована совокупностью нейронов разных размеров и форм, распределенных как диффузно, так и в виде ядер. Нейроны ретикулярной формации имеют мало ветвящиеся дендриты и хорошо ветвящиеся аксоны, которые часто образуют Т-образное ветвление: одна из ветвей аксона имеет нисходящее, а другое – восходящее направление. Ветви нейронов имеют под микроскопом вид сеточки (ретикулума), с чем связано название данной структуры мозга.
К наиболее крупным ядрам ретикулярной формации на уровне продолговатого мозга относятся: гигантоклеточное ретикулярное, мелкоклеточное ретикулярное, латеральное ретикулярное ядро. На уровне моста – каудальное, вентрикулярное и оральное ретикулярные ядра.
Нейроны ретикулярной формации характеризуются богатством связей. От них начинаются как восходящие, так и нисходящие пути, дающие многочисленные коллатерали, образующие синаптические контакты на нейронах ядер ЦНС (ядра черепно-мозговых нервов, мозжечка, промежуточного мозга, коры больших полушарий).
Афферентные входы поступают в ретикулярную формацию преимущественно от трех источников:
1) от температурных и болевых рецепторов по волокнам спиноретикулярного пути и тройничного нерва;
2) от сенсорной и частично от других зон коры головного мозга по кортико-ретикулярным путям импульсация поступает в ядра, дающие начало ретикулоспинальным путям, а также в ядра, которые проецируются на мозжечок;
3) от ядер мозжечка по мозжечковоретикулярному пути.
Эфферентные выходы из ретикулярной формации проецируются:
1) в спинной мозг по ретикулоспинальному тракту;
2) к верхним отделам головного мозга;
Многообразие связей и структур ретикулярной формации определяет ее многочисленные функции, которые можно объединить в три главные группы: соматические, сенсорные и вегетативные. Вместе с тем ретикулярная формация обеспечивает регуляцию деятельности различных отделов ЦНС.
Соматические функции ретикулярной формации проявляются в ее влиянии на двигательные ядра черепно-мозговых нервов, моторные спинальные центры и активность мышечных рецепторов (мышечных веретен).
В парамедианной ретикулярной формации продолговатого мозга имеются нейроны, управляющие горизонтальными движениями глаз и опосредованно, через нейроны ретикулярной формации среднего мозга, управляют вертикальными движениями глаз. При двусторонних поражениях этих отделов ретикулярной формации становится невозможным произвольное выполнение горизонтальных и вертикальных движений глаз.
Через латеральный ретикулоспинальный путь, волокна которого оканчиваются на вставочных нейронах спинного мозга, происходит возбуждение альфа- и гамма-нейронов мышц-сгибателей конечностей и реципрокное торможение мышц-разгибателей. Латеральный ретикулоспинальный путь совместно с руброспинальным и кортикоспинальным путями, сходным с ним в функциональном и топографическом плане, образуют нисходящую латеральную флексорную (сгибательную) систему.
Раздражение нейронов центральной части гигантоклеточного ретикулярного ядра в эксперименте вызывает подавление всех спинальных рефлексов (сгибательных и разгибательных). По этому механизму осуществляется открытое И.М. Сеченовым (1863) центральное торможение спинного мозга.
По медиальному ретикулоспинальному пути, идущему от ретикулярных ядер моста, осуществляется стимуляция альфа- и гамма-нейронов мышц-разгибателей осевой мускулатуры тела (туловища и проксимальных отделов конечностей) и через тормозные интернейроны тормозятся сгибатели. Этот путь совместно с вестибулоспинальными путями составляет медиальную нисходящую экстензорную (разгибательную) систему.
Восходящее влияние ретикулярной формации на кору головного мозга. Ретикулярная формация регулирует уровень возбудимости и тонуса различных отделов нервной системы, включая кору больших полушарий.
Еще в 1935 г. Ф. Бремером в остром опыте было показано, что при перерезке ствола мозга между верхними и нижними буграми четверохолмия (препарат cervea isole) животное впадало в сноподобное состояние: отсутствовала реакция на световые и обонятельные раздражители.
На электроэнцефалограмме (ЭЭГ) у таких животных преобладал регулярный низкочастотный, высокоамплитудный альфа-ритм, который свидетельствовал о синхронизации биоритмов структур коры большого мозга.
У человека альфа-ритм преобладает в ЭЭГ при состоянии покоя и отсутствии внешних раздражений.
В хронических опытах с электрической стимуляцией ретикулярной формации X. Мэгуном и Дж. Моруцци в 1949 г. были установлены ее восходящие, активирующие влияния на кору головного мозга. Было показано, что при раздражении центральной части ретикулярной формации сонная кошка пробуждалась и у нее возникала ориентировочная реакция. На электроэнцефалограмме (ЭЭГ) пробуждение животного проявлялось в смене альфа-ритма низкоамплитудным, высокочастотным бета-ритмом. Данная электроэнцефалографическая реакция получила название реакции десинхронизации. Она имеет генерализованный характер и регистрируется в различных зонах коры головного мозга.
Восходящее влияние ретикулярной формации на большой мозг может быть и тормозным. По данным Дж. Моруцци, раздражение некоторых участков ретикулярной формации заднего мозга может вызвать у животного глубокий сон и появление альфа-ритма в электроэнцефалограмме.
На основании этих экспериментальных данных сложилось представление, что важнейшей функцией восходящей ретикулярной формации является регуляция цикла сон–бодрствование и уровня сознания.
Дыхательный центр находится в области IV мозгового желудочка. Повреждение этой области приводит к остановке дыхания. Нейроны дыхательного центра обладают способностью к автоматии, т. е. периодической активности в отсутствии афферентных воздействий.
В дыхательном центре выделены две зоны: инспираторная (дорсальная), связанная с осуществлением акта вдоха, и экспираторная (вентральная) – с актом выдоха.
Дорсальная зона включает в себя большую часть ретикулярного гигантоклеточного ядра, состоит на 95 % из нейронов вдоха. Отростки этих нейронов в составе ретикулоспинальных путей иннервируют ядра диафрагмальных нервов. Аксоны небольшой части инспираторных нейронов вентральной зоны иннервируют мотонейроны ядер межреберных мышц и частично ядра диафрагмальных нервов. Импульсация в инспираторных дорсальных нейронах начинается за 0,1 сек до вдоха сначала в нейронах дорсальной, а затем в нейронах вентральной зоны. Поэтому диафрагма начинает сокращаться раньше, чем наружные межреберные мышцы. В вентральной зоне находятся преимущественно нейроны выдоха. Аксоны этих нейронов иннервируют мотонейроны внутренних межреберных и брюшных мышц. Импульсация в мотонейронах появляется за 0,1 сек до выдоха, затем частота ее нарастает в течение выдоха и резко уменьшается в конце.
К регуляции дыхания имеет также отношение пневмотаксический центр, который находится в верхней области моста, а также апнейстический центр, нейроны которого располагаются в средней и нижней частях моста. Пневмотаксический центр ограничивает длительность вдоха и повышает частоту дыхания. Эффект действия апнейстического центра заключается в постоянной стимуляции инспираторных нейронов, он способствует возникновению вдоха и удлиняет его.
Смена фаз дыхательного цикла может осуществляться рефлекторно за счет афферерентных влияний, которые модулируют дыхательную ритмику. При интенсивном вдохе и растяжении легочной ткани импульсы от механорецепторов легких по афферентным волокнам блуждающего нерва приходят в дыхательный центр и вызывают рефлекторное торможение инспираторных нейронов, одновременно возбуждая экспираторные (рефлекс Геринга–Брейера). Импульсы от экспираторных нейронов по ретикулоспинальному пути достигают исполнительных моторных центров спинного мозга, стимулируют начало выдоха.
Важной особенностью дыхательного центра является то, что он активируется не только афферентными импульсами, но и химическими раздражителями. Произвольная регуляция дыхания осуществляется корой больших полушарий.
Другим жизненно важным центром ретикулярной формации является сердечно-сосудистый центр. Его локализация впервые была определена Ф.В. Овсянниковым (1871 г.). Он занимает обширную область продолговатого мозга, простираясь в дорсолатеральном направлении от дна IV желудочка до пирамид.
Рефлекторные влияния на нейроны сосудодвигательного центра осуществляются при возбуждении хемо- и механорецепторов, локализованных в сосудистой стенке. Афферентные волокна от этих рецепторов в составе блуждающего и языкоглоточного нервов достигают продолговатого мозга. Возбуждение механорецепторов дуги аорты, каротидного синуса при повышении артериального давления вызывает торможение активности сосудодвигательного центра и, как следствие, рефлекторное снижение сосудистого тонуса (рефлексы Людвига–Циона, Геринга, Бейнбриджа). Напротив, при повышении давления в системе полых вен наблюдается усиление активности сосудистого центра и вазоконстрикторный эффект. Тонус сосудистого центра может изменяться и при возбуждении хеморецепторов сосудистой стенки, возникающем при изменениях химического состава крови.
Следует отметить, что деятельность сосудодвигательного центра сочетается с функцией моторного ядра блуждающего нерва – двойного ядра, снижающего в норме частоту сердечных сокращений. В связи с этим при вазоконстрикторном эффекте одновременно увеличивается частота сердечных сокращений и, наоборот, при вазодилатации наблюдается замедление сердечной ритмики.
Что делают нейроны ретикулярной формации продолговатого мозга
Разнообразие функций, осуществляемых различными отделами ретикулярной формации, представлено в таблице ниже.
а) Генераторы программ движений. К программам движений черепных нервов относят следующие:
• Содружественные (параллельные) движения глаз, местно контролируемые двигательными узлами (центрами взора) в среднем мозге и мосту, имеющие связь с ядрами двигательных нервов глаз.
• Ритмичные жевательные движения, контролируемые супратригеминальным премоторным ядром моста.
• Глотание, рвотные движения, кашель, зевание и чихание контролируют отдельные премоторные ядра продолговатого мозга, имеющие связь с соответствующими черепными нервами и дыхательным центром.
Слюноотделительные ядра относят к мелкоклеточной ретикулярной формации моста и продолговатого мозга. От них отходят преганглионарные парасимпатические волокна к лицевому и языкоглоточному нервам.
 Ретикулярная формация (РФ).
Ретикулярная формация (РФ).
(А) Отделы. (Б) Группы аминергических и холинергических клеток. 
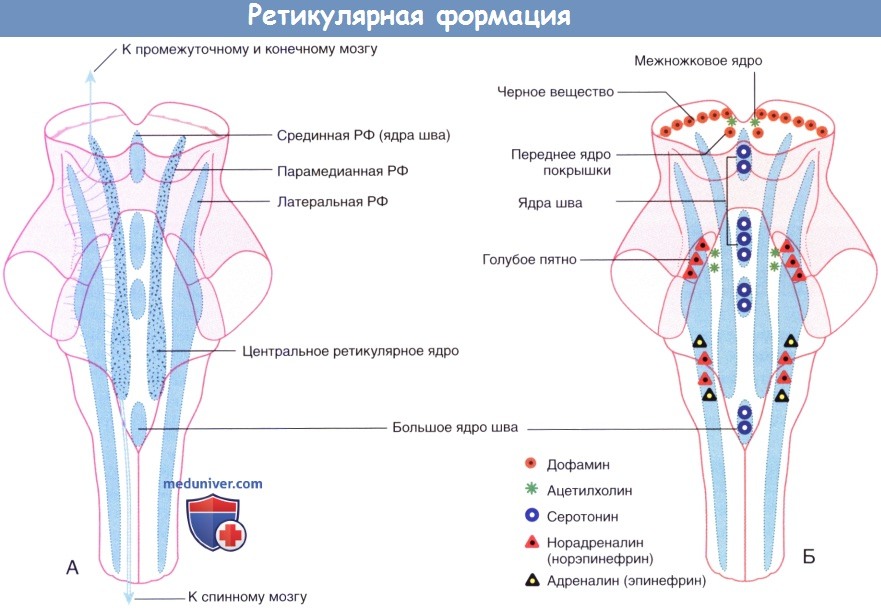
1. Генераторы программ движений. Из экспериментов на животных давно установлено, что генераторы программ движений низших позвоночных и низших млекопитающих расположены в сером веществе спинного мозга, соединяясь с помощью нервов с каждой из четырех конечностей. Данные генераторы в спинном мозге представляют собой электрические нейронные сети, последовательно доставляющие сигналы к сгибательным и разгибательным группам мышц. Генераторная активность спинного мозга подчиняется командам из высших центров—двигательной области среднего мозга (ДОСМ).
К ДОСМ относят ножкомостовое ядро, прилежащее к верхней мозжечковой ножке в месте ее прохождения в области верхнего края четвертого желудочка и соединения со средним мозгом. От этих ядер в составе центрального покрышечного пути отходят нисходящие волокна к оральному и каудальному ядрам моста, образованным двигательными нейронами, иннервирующими мышцы разгибатели, и к крупноклеточным нейронам продолговатого мозга, контролирующим нейроны, иннервирующие мышцы-сгибатели.
Основной механизм реабилитации при поражениях спинного мозга—активация спинномозговых двигательных рефлексов у пациентов, перенесших травмы с частичным или полным разрывом спинного мозга. В настоящее время хорошо известно, что даже после полного разрыва на уровне шейного или грудного отдела возможна активация пояснично-крестцовых программ движений путем продолжительной электрической стимуляции твердой мозговой оболочки на уровне поясничных сегментов. Стимуляция в значительной степени активирует волокна задних корешков, запуская образование импульсов в основании переднего рога.
При поверхностной электромиографии (ЭМГ) со сгибательных и разгибательных мышц было обнаружено последовательное возбуждение нейронов мышц сгибателей и разгибателей, хотя данная программа не соответствовала нормальной. Для образования нормальной программы разрыв должен быть неполным с сохранением части нисходящих путей от ножкомостового ядра.
Создание истинных шагательных движений при полном разрыве возможно, если пациента поставить на беговую дорожку с одновременной стимуляцией твердой мозговой оболочки, преимущественно за счет получения генератором дополнительных чувствительных и проприоцептивных импульсов. Сила мышц и скорость ходьбы будут нарастать в течение нескольких недель, но недостаточно для ходьбы без использования ходунков.
Современные исследования направлены на улучшение возможности «создания моста» с супраспинальными двигательными волокнами путем очищения от остатков тканей в месте разрыва и замещения этих тканей составом, физически и химически стимулирующим регенерацию аксонов.
2. Высшие центры контроля мочеиспускания описаны в следующей статье на сайте.
 Общая схема контроля движений.
Общая схема контроля движений.
б) Контроль дыхания. Дыхательный цикл в значительной степени регулируют дорсальное и вентральное дыхательные ядра, расположенные в верхнем отделе продолговатого мозга с каждой стороны от срединной линии. Дорзальное дыхательное ядро расположено в среднелатеральном отделе ядра одиночного пути. Вентральное ядро расположено позади двойного ядра (отсюда название — позадидвойное ядро). Оно отвечает за выдох; поскольку этот процесс в норме происходит пассивно, активность нейронов при обычном дыхании относительно низкая, но значительно возрастает при нагрузках. Третье, медиальное парабрахиальное ядро, прилежащее к голубому пятну, вероятно, имеет значение в механизме дыхания, возникающем в состоянии бодрствования.
Парабрахиальное ядро, образованное множеством подгрупп нейронов, вместе с описанными выше аминергической и холинергической системами участвует в поддержании состояния бодрствования путем активации коры головного мозга. Стимуляция этого ядра миндалевидным телом при тревожных расстройствах приводит к характерной гипервентиляции.
Дорзальное дыхательное ядро контролирует процесс вдоха. От него отходят волокна к двигательным нейронам на противоположной стороне спинного мозга, иннервирующим диафрагму, межреберные и вспомогательные дыхательные мышцы. Ядро получает восходящие возбуждающие импульсы от хеморецепторов хемочувствительной области продолговатого мозга и каротидного синуса.
Вентральное дыхательное ядро отвечает за выдох. При спокойном дыхании оно работает как нейронная цепь, участвуя в реципрокном торможении центра вдоха посредством ГАМК-ергических (γ-аминомасляная кислота) вставочных нейронов. При форсированном дыхании оно активирует клетки переднего рога, иннервирующие брюшные мышцы, отвечающие за спадение легких.
1. Хемочувствительная область продолговатого мозга. Сосудистое сплетение четвертого желудочка продуцирует спинномозговую жидкость (СМЖ), проходящую через латеральную апертуру (Лушка) четвертого желудочка. Клетки латеральной ретикулярной формации на поверхности продолговатого мозга в этой области имеют исключительную чувствительность к концентрации ионов водорода (Н + ) в омывающей СМЖ. Фактически, эта хемочувствительная область продолговатого мозга анализирует парциальное давление углекислого газа (рСО2) в СМЖ, которое соответствует рСО2 крови, снабжающей головной мозг. Любое повышение концентрации ионов Н + приводит к стимуляции дорсального дыхательного ядра путем прямой синаптической связи (в продолговатом мозге расположено несколько других хемочувствительных ядер).
2. Хеморецепторы каротидного синуса. Каротидный синус размером с булавочную головку прилежит к стволу внутренней сонной артерии и получает от этой артерии веточку, разветвляющуюся внутри. Кровоток через каротидный синус настолько интенсивный, что артериовенозное парциальное давление кислорода (рО2) изменяется менее чем на 1 %. Хеморецепторы представляют собой клетки клубочка, иннервируемые ветвями синусного нерва (ветвь IX черепного нерва). Каротидные хеморецепторы реагируют как на снижение рО2, так и на повышение рСО2 и обеспечивают рефлекторную регуляцию уровней газов крови путем изменения частоты дыхания.
Хеморецепторы аортального гломуса (под дугой аорты) у человека относительно недоразвиты.
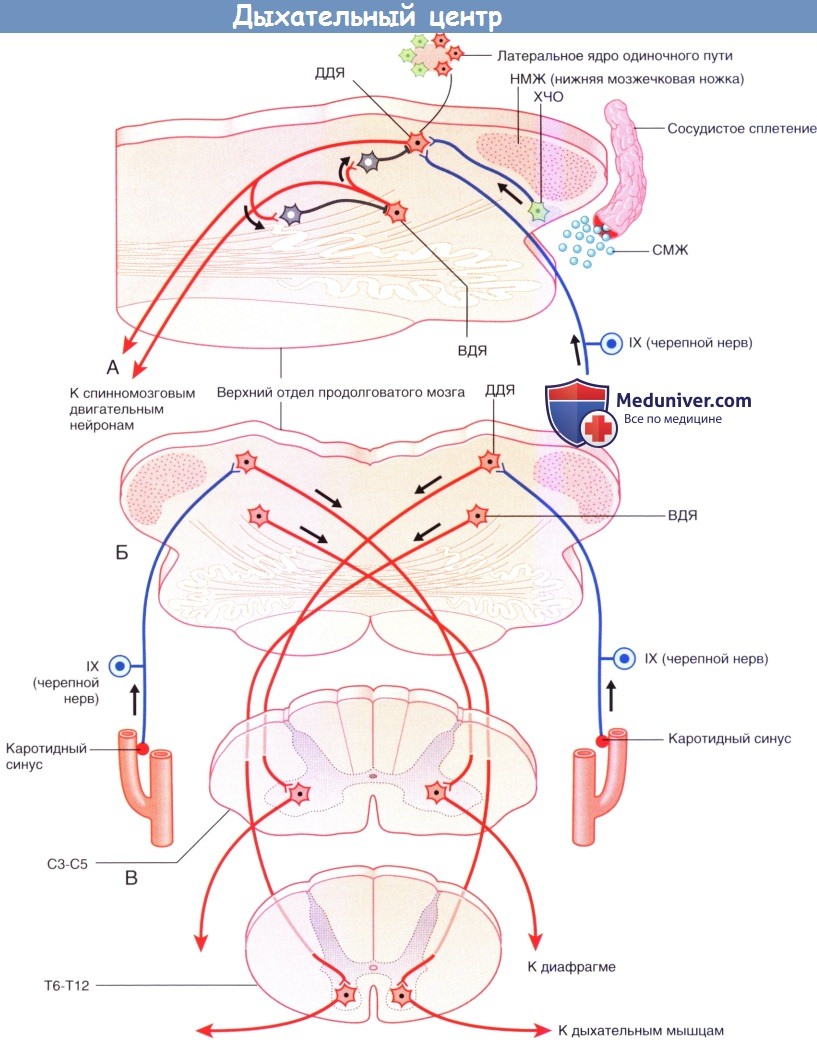 Дыхательный центр. Все срезы показаны снизу и сзади.
Дыхательный центр. Все срезы показаны снизу и сзади.
(А) — увеличенный срез (Б).
(А) Тормозное взаимодействие между дорсальным и вентральным дыхательными ядрами (ДДЯ, ВДЯ).
К хемочувствительной области (ХЧО) продолговатого мозга, волокна от которой направляются к ДДЯ, прилежат капилляры сосудистой оболочки, продуцирующие спинномозговую жидкость (СМЖ) (Б).
В составе языкоглоточного нерва (IX) проходят хемочувствительные волокна от каротидного синуса до ДДЯ.
(В) Возбуждение двигательных нейронов диафрагмы осуществляет противоположное ДДЯ.
(Г) Для форсированного выдоха ВДЯ противоположной стороны возбуждает нейроны мышц передней брюшной стенки.
в) Контроль сердечно-сосудистой системы. Сердечный выброс и периферическое сопротивление сосудов регулируют нервная и эндокринная системы. Вследствие широкого распространения эссенциальной артериальной гипертензии в позднем среднем возрасте большинство исследований в этой области направлено на изучение механизмов сердечно-сосудистой регуляции.
Восходящие волокна, сигнализирующие о повышенном артериальном давлении, начинаются от рецепторов растяжения (многочисленных свободных нервных окончаний) в стенке каротидного синуса и дуги аорты. Эти восходящие волокна, известные как барорецепторы, направляются к медиально расположенным клеткам ядра одиночного пути, образующим барорецепторный центр. Восходящие волокна от каротидного синуса проходят в составе языкоглоточного нерва; волокна от дуги аорты входят в состав блуждающего нерва. Барорецепторные нервы относят к «буферным нервам», так как их действие заключается в коррекции любых отклонений артериального давления от нормы.
Сердечный выброс и периферическое сопротивление сосудов зависят от активности симпатической и парасимпатической нервных систем. Два основных барорецепторных рефлекса — парасимпатический и симпатический — способствуют нормализации повышенного артериального давления.
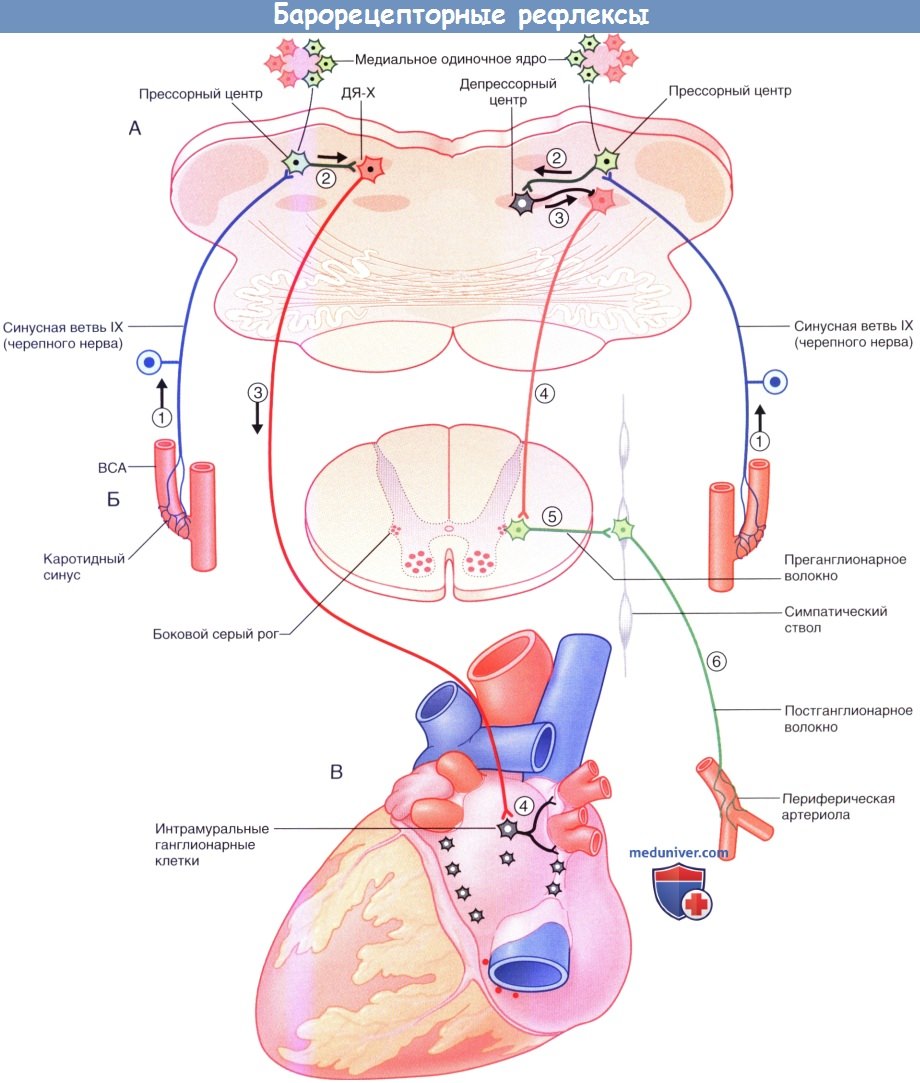 Барорецепторные рефлексы:
Барорецепторные рефлексы:
(А) Верхний отдел продолговатого мозга.
(Б) Сегменты спинного мозга от Т1 до L3.
(В) Задняя стенка сердца. Барорецепторный рефлекс (слева):
1. Рецепторы растяжения в каротидном синусе возбуждают волокна синусовой ветви языкоглоточного нерва. ВСА—внутренняя сонная артерия.
2. Барорецепторные нейроны ядра одиночного пути отвечают возбуждением тормозящих сердце (кардиоингибирующих) нейронов дорсального (двигательного) ядра блуждающего нерва (ДЯ-Х).
3. Преганглионарные парасимпатические холинергические волокна блуждающего нерва образуют синапсы с клетками интрамуральных ганглиев в задней стенке сердца.
4. Постгангионарные парасимпатические холинергические волокна тормозят пейсмекерную активность синоатриального узла, уменьшая, тем самым, частоту сердечных сокращений.
Баросимпатический рефлекс (справа):
1 Афферентные волокна рецепторов растяжения каротидного синуса возбуждают медиальные барорецепторные нейроны ядра одиночного пути.
2. Барорецепторные нейроны отвечают возбуждением тормозных нейронов депрессорного центра в центральном ретикулярном ядре продолговатого мозга.
3. Происходит торможение адренергических и норадренергических нейронов прессорного центра латерального ретикулярного ядра (переднего вентролатерального отдела продолговатого мозга).
4. Уменьшается тоническое возбуждение нейронов боковых рогов спинного мозга.
5 и 6. Происходит пре- и постганглионарное торможение симпатической иннервации тонуса артериол, что, в свою очередь, приводит к снижению периферического сосудистого сопротивления.
г) Сон и бодрствование. При электроэнцефалографии (ЭЭГ) можно наблюдать характерные картины электрической активности корковых нейронов при разных состояниях сознания. Нормальное состояние бодрствования характеризуется высокочастотными низкоамплитудными волнами. Погружение в сон сопровождается низкочастотными высокоамплитудными волнами, более высокая амплитуда волн обусловлена синхронизированной активностью большего числа нейронов. Такой тип сна называют медленноволновым (синхронизированным), или He-REM-сном (REM—rapid eye movement— быстрые движения глаз). Он продолжается около 60 мин, а затем переходит в десинхронизированный сон, при котором последовательности на ЭЭГ напоминают таковые при состоянии бодрствования. Только в этот период возникают сны и быстрые движения глаз (отсюда и более часто употребляемый термин — REM-сон). В период нормального ночного сна сменяют друг друга несколько циклов REM-сна и He-REM-сна, описанные в отдельной статье на сайте.
Смена циклов сна и бодрствования — отражение двух нейронных сетей мозга, одной — работающей в состоянии бодрствования, а другой — в состоянии сна. Эти сети проти вопоставлены друг другу по типу «переключателя» между сном и бодрствованием (что делает возможным переключение между сетями быстрым и полным). Аналогичная схема работает при смене REM-сна на медленноволновой сон. В норме управление сном происходит с помощью физиологических систем (вклад системы гомеостаза — изменение уровня метаболизма клеток), циркадных ритмов (супрахиазмальное ядро— главные биологические часы, которые синхронизированы с информацией от окружающей среды, воздействием света на сетчатку и мелатонином, вырабатываемым эпифизом, и управляют циклом сон-бодрствование и другими физиологическими функциями) и аллостатической нагрузки (принятие пищи и двигательная активность).
Эти факторы изменяются медленно, и без быстрой смены состояния переключательного механизма переход от бодрствования ко сну также был бы медленным и неудобным.
3. Стимуляция пробуждения, или активирующие системы (каудальный отдел среднего мозга и ростральный отдел моста). За активацию коры больших полушарий отвечают два основных пути:
• Холинергические нейроны (ножкомостового и латеродорсальных ядер покрышки) подходят к таламусу (переключательным ядрам и ретикулярному ядру) и ингибируют те ГАМК-ергические нейроны таламуса, задача которых — препятствование передаче чувствительной информации к коре полушарий.
• Моноаминергические нейроны расположены в голубом пятне, дорсальном и срединном ядрах шва (серотонинергические), парабрахиальном ядре (глутаматергические), околоводопроводном сером веществе (ОВСВ, дофаминергические) и в серобугорно-сосцевидном ядре (гистаминергические). Аксоны нейронов каждой из этих областей направляются к базальным отделам переднего мозга (базальному ядру Мейнерта и безымянной субстанции), а оттуда — к коре больших полушарий.
Пептидергические (орексиновые) и глутаматергические нейроны латерального гипоталамуса, а также холинергические и ГАМК-ергические нейроны базальных ганглиев переднего мозга также посылают волокна к коре больших полушарий.
4. Система индукции сна (гипоталамус). Нейроны вентролатерального преоптического ядра (продуцирующие ГАМК и галанин — тормозные нейромедиаторы) иннервируют большую часть отделов активирующих систем; они активируются в основном во время сна.
Нейроны срединного преоптического ядра—также ГАМК-ергические. Считают, что они отвечают на изменения гомеостаза (указывающие на накопившуюся потребность во сне) и активируют нейроны вентролатерального преоптического ядра.
Нейроны каудолатеральной мостовой ретикулярной формации — также ГАМК-ергические, их восходящие аксоны оказывают сон-индуцирующее воздействие.
5. Переключательный механизм между сном и бодрствованием. Во сне ГАМК-ергические нейроны системы индукции сна (вентролатеральное преоптическое и срединное преоптическое ядра) активно тормозят холинергические и моноаминергические нейроны восходящей активирующей системы путем выделения ГАМК. Противоположный результат возникает в случае, когда восходящая активирующая система тормозит сон-индуцирующие нейронные цепи, поддерживая состояние бодрствования.
6. Центры REM-сна и He-REM-сна (мост). Нейроны верхнего отдела моста (области подголубого пятна и сублатеродорсального ядра) образуют «центр REM-сна» и отвечают за наступление REM-сна. Восходящие пути от разных групп этих нейронов направляются к гипоталамусу и базальным ядрам переднего мозга (образующим ЭЭГ-картины REM-сна и сновидения), а нисходящие пути — к стволу мозга (отвечает за быстрые движения глаз, снижение тонуса мышц и торможение двигательной активности путем стимуляции вставочных тормозных нейронов спинного мозга, приводящих к гиперполяризации а-мотонейронов).
Центр REM-сна в норме ингибирован рядом расположенными ГАМК-ергическими вставочными нейронами (лежащими кпереди от вентролатерального отдела ОВСВ и прилежащей части латерального отдела покрышки моста), а также нейронами центра He-REM-сна. При активности этих нейронов период REM-сна не наступает. Если возбуждены нейроны центра REM-сна, происходит торможение центра He-REM-сна (переключательный механизм между REM- и He-REM-сном), что способствует быстрому переходу между REM- и He-REM-сном.
Центр He-REM-сна получает волокна от множества отделов мозга и участвует в регуляции REM-сна. Возбуждающие импульсы от гипоталамуса (орексиновые нейроны) и моста (моноаминергические из голубого ядра и дорсальных ядер шва) приводят к подавлению REM-сна, тогда как тормозные волокна из гипоталамуса (ГАМК-ергические нейроны вентролатерального преоптического ядра и холинергические латеродорсального и ножкомостового ядер покрышки) стимулируют переход к REM-сну.
Большое разнообразие расстройств сна можно объяснить нарушением системы сон-бодрствование и КЕМ-не-КЕМ-систем.
7. Восходящая активирующая система. В более ранних исследованиях были выявлены нейроны ретикулярной формации ствола мозга (от среднего мозга до продолговатого мозга), играющие роль в регуляции сна и бодрствования. Такое анатомическое расположение, важность их роли и широкое воздействие на весь мозг и привели к появлению названия восходящая активирующая ретикулярная система (ВАРС). Однако в настоящее время стало известно, что к этой системе относят не только ретикулярную формацию, поэтому более предпочтительный термин — восходящая активирующая система.
Основную часть этой системы составляет голубое пятно (норадренергическое), дорсальное и срединное ядра шва (серотонинергические), ножкомостовое и латеродорсальное ядра покрышки (холинергические), серобугорно-сосцевидное ядро (гистаминергическое). К этой системе также относят гипоталамус (орексинергические нейроны), базальные ядра переднего мозга и другие группы нейронов. Активация поддерживается за счет влияния вышеперечисленных групп нейронов на таламус и кору больших полушарий, а также их связи с циклом сон-бодрствование. Их общий вклад в цикл сон-бодрствование частично избыточен, поэтому повреждение одного из отделов обычно не приводит к полному нарушению состояния бодрствования.
д) Сенсорная модуляция: воротный контроль. Передача чувствительной информации с первичных афферентных нейронов на вторичные (на уровне задних рогов спинного мозга и дорсальных ядер) и от вторичных нейронов на третичные афферентные нейроны (на уровне таламуса) — часть воротного механизма. Термин воротный механизм указывает на степень свободы передачи импульсов с одной группы нейронов на следующую.
Воротный контроль передачи тактильной чувствительности происходит на уровне дорсальных ядер спинного мозга. Кортико-спинномозговые волокна от нейронов постцентральной извилины облегчают или тормозят на этом уровне передачу чувствительной информации, что подробно описано в главе 16.
Регуляция передачи ноцицептивной информации от туловища и нижних конечностей происходит в задних рогах спинного мозга. Контроль передачи чувствительности от головы и верхней половины тела осуществляется в спинномозговом ядре тройничного нерва. Ключевое образование заднего рога — желатинозная субстанция, заполненная мелкими возбуждающими и тормозными вставочными нейронами. Возбуждающий нейромедиатор — глутамат, тормозной медиатор — ГАМК в одной группе вставочных нейронов и энкефалин (опиоидный пентапептид) — в другой.
Высокомиелинизированные (Аδ) полимодальные ноцицептивные нервные волокна контактируют непосредственно с дендритами переключающих (релейных) нейронов, образующих спиноталамический тракт, и с аналогичными нейронами тройничного ядра. По Аδ-волокнам происходит передача острой, хорошо локализованной боли. Безмиелиновые ноцицептивные афферентные волокна С-типа не имеют прямого переключения на релейные клетки через вставочные нейроны в желатинозной субстанции. По волокнам С-типа происходит передача тупой, плохо локализованной боли. Большая их часть содержит субстанцию Р, которая играет роль вторичного нейромедиатора для глутамата.
1. Сегментарная антиноцицептивная система. Крупные (A-типа) механорецепторные афферентные волокна от волосяных фолликулов образуют синапсы со спиноталамическими переключательными нейронами и их аналогами в ядре тройничного нерва. Они также отдают коллатеральные волокна к тормозным (преимущественно ГАМК-ергическим) клеткам желатинозной субстанции, которые, в свою очередь, образуют синапсы со спиноталамическими переключательными нейронами. Часть вставочных нейронов оказывает пресинаптическое торможение терминальных волокон С-типа через аксоаксональиые контакты (которые крайне трудно обнаружить в эксперименте) или аксодентритические контакты. Проведение сниноталамических импульсов в ответ на возбуждение С-волокна может быть усилено путем стимуляции афферентных волокон от механорецепторов, что приводит к возбуждению тормозных желатинозных клеток.
Эта простая нервная цепочка приводит к облегчению боли путем «растирания болезненного участка». Она также объясняет целесообразность использования метода чрескожной электростимуляции нервов (ЧЭСН) физиотерапевтами для облегчения боли при артрите и других хронических болевых расстройствах. При стандартной методике ЧЭСН стимулирующий электрод накладывают на участок кожи на том же сегментарном уровне, где находится источник болевого раздражения, проходящего по волокнам С-типа, и пропускают ток с силой, достаточной для появления сильного зуда.
2. Супраспинальный уровень антиноцицептивной системы. Большое ядро шва. От большого ядра шва (БЯШ) продолговатого мозга в составе пучка Лиссауэра нисходят двусторонние шовно-спинномозговые волокна, оканчивающиеся в желатинозной субстанции на всех уровнях спинного мозга. Электростимуляция БЯШ у животных может приводить к полной аналгезии всего тела, оказывая лишь небольшое влияние на тактильную чувствительность. Медиатором большой части волокон выступает серотонин, возбуждающий тормозные вставочные нейроны в задних рогах и спинномозговом ядре тройничного нерва. Вставочные нейроны отвечают за пре- и постсинаптическое торможение соответствующих релейных клеток.
Имеются данные о том, что норадренергические волокна, проходящие от моста и продолговатого мозга к задним рогам, также принимают участие в антиноцицептивной системе на супраспинальном уровне путем прямого торможения спиноталамических нейронов.
• Диффузный тормозной контроль боли. БЯШ не имеет соматотопической организации, однако оно получает информацию от спиноретикулярных и тригеминоретикулярных нейронов, отвечающих за периферическую болевую стимуляцию. Такую анатомическую связь обозначают термином «диффузный тормозной контроль боли». Болевая стимуляция в одном участке тела может облегчить боль в других его участках. Подобное строение хорошо объясняет гетеротопическое ослабление боли при акупунктуре, когда иглы используют для возбуждения восходящих болевых волокон в наиболее поверхностно расположенных мышцах, а не в коже.
• Стимул-индуцированная аналгезия. БЯШ непосредственно отвечает за стимуляцию ОВСВ среднего мозга. Эту связь используют для помощи пациентам с хроническим болевым синдромом: небольшой стимулирующий электрод устанавливают в ОВСВ, а пациенту позволяют контролировать уровень стимуляции.
• Стресс-индуцированная аналгезия. В состоянии покоя волокна ОВСВ к БЯШ находятся в состоянии тонического торможения вставочными (ГАМК-ергическими) нейронами, расположенными в ОВСВ. Вставочные нейроны, в свою очередь, ингибированы опиоидными пептидами (и каннабиноидами) — в значительной степени β-эндорфином, высвобождаемым небольшой группой гипоталамических нейронов, волокна которых направляются к ОВСВ. При жизнеугрожающих ситуациях, когда травма становится необходимой для спасения, ОВСВ может быть расторможено гипоталамусом. Вероятно, существует механизм, на основании которого огнестрельная рана может быть едва замечена в пылу битвы (как будет описано в отдельной статье на сайте, возбуждающие нейроны ОВСВ во время страха также могут активироваться непосредственно миндалевидным телом, расположенным в передней височной доле).
Помимо сегментарного и супраспинального уровней контроля ноцицептивной передачи от первичных нейронов к вторичным, воротный контроль также имеется в таламусе.
Кроме того, компонент отвращения (неприятного ощущения) при боли требует участия передней поясной коры, богатой опиоидными рецепторами.
е) Резюме. Ретикулярная формация расположена на всем протяжении ствола мозга, преимущественно в виде трех клеточных рядов. Латеральная (мелкоклеточная) группа клеток получает восходящие волокна от чувствительных ядер всех черепных и периферических нервов. От нее отходят волокна в парамедианную крупноклеточную ретикулярную формацию, аксоны которой, в свою очередь, направляются в головной и спинной мозг. Срединная ретикулярная формация образована серотонинергическими нейронами.
1. Аминергические нейроны. Волокна от серотонинергических нейронов ядер шва направляются во все отделы серого вещества ЦНС. Волокна дофаминергических нейронов проходят от черного вещества к полосатому телу, а также от вентральных ядер покрышки среднего мозга к лобной коре и добавочному ядру. Норадренергические нейроны голубого пятна посылают волокна ко всем отделам серого вещества ЦНС. Волокна адренергических нейронов продолговатого мозга идут в гипоталамус и спинной мозг.
2. Генераторы программ движений. Центры взора среднего мозга и моста образуют содружественные движения глаз; локомоторный центр среднего мозга отвечает за ходьбу; супратригеминальное ядро моста регулирует процесс жевания; мостовой центр контроля мочеиспускания контролирует деятельность мочевого пузыря. В продолговатом мозге расположены дыхательный, рвотный, кашлевой центры и центр чихания, а также прессорный и депрессорный отделы сосудодвигательного центра. Кроме того, в хемочувствительной области продолговатого мозга расположены нейроны ретикулярной формации, чувствительные к уровню ионов H + в СМЖ.
Сон и бодрствование регулируют серотонин- и норадренергические нейроны, а также холинергические нейроны верхнего отдела моста. Восходящая активирующая система—анатомически-физиологическое образование, включающее группы нейронов ствола мозга, оказывающих возбуждающее воздействие на мозг, что можно зарегистрировать при ЭЭГ. Это воздействие опосредовано влиянием на таламус и кору полушарий.
3. Антиноцицептивная система. Активация сегментарной антиноцицептивной системы происходит путем стимуляции волокон A-типа от волосяных фолликулов. Супраспинальная антиноцицептивная система—функция БЯШ продолговатого мозга, которое возбуждают гипоталамус и средний мозг. Серотонин из терминалей БЯШ в желатинозной субстанции задних рогов спинного мозга и спинномозговом ядре тройничного нерва стимулирует энкефалинергические вставочные нейроны, тормозящие передачу импульсов на спиноталамические/тригемино-таламические нейроны.
Редактор: Искандер Милевски. Дата публикации: 20.11.2018