Ультразвуковое исследование мозга новорожденных детей (нормальная анатомия)


Журнал «SonoAce Ultrasound»
Содержит актуальную клиническую информацию по ультрасонографии и ориентирован на врачей ультразвуковой диагностики, выходит с 1996 года.
Показания для проведения эхографии мозга
Акустическим окном для исследования мозга может служить любое естественное отверстие в черепе, но в большинстве случаев используют большой родничок, поскольку он наиболее крупный и закрывается последним. Маленький размер родничка значительно ограничивает поле зрения, особенно при оценке периферических отделов мозга.
Для проведения эхоэнцефалографического исследования датчик располагают над передним родничком, ориентируя его так, чтобы получить ряд корональных (фронтальных) срезов, после чего переворачивают на 90° для выполнения сагиттального и парасагиттального сканирования. К дополнительным подходам относят сканирование через височную кость над ушной раковиной (аксиальный срез), а также сканирование через открытые швы, задний родничок и область атланто-затылочного сочленения.
По своей эхогенности структуры мозга и черепа могут быть разделены на три категории:
Нормальные варианты мозговых структур
Борозды и извилины. Борозды выглядят как эхогенные линейные структуры, разделяющие извилины. Активная дифференцировка извилин начинается с 28-й недели гестации; их анатомическое появление предшествует эхографической визуализации на 2-6 нед. Таким образом, по количеству и степени выраженности борозд можно судить о гестационном возрасте ребенка.
Сосудистые сплетения могут быть источником внутрижелудочковых кровоизлияний у доношенных детей, тогда на эхограммах видна их четкая асимметрия и локальные уплотнения, на месте которых затем образуются кисты.
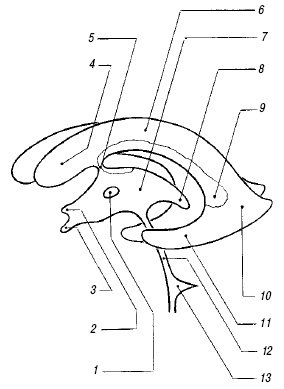
Сильвиев водопровод и IV желудочек. Сильвиев водопровод (aquaeductus cerebri) представляет собой тонкий канал, соединяющий III и IV желудочки (см. рис. 1), редко видимый при УЗ исследовании в стандартных позициях. Его можно визуализировать на аксиальном срезе в виде двух эхогенных точек на фоне гипоэхогенных ножек мозга.
IV желудочек (ventriculus quartus) представляет собой небольшую полость ромбовидной формы. На эхограммах в строго сагиттальном срезе он выглядит малым анэхогенным треугольником посередине эхогенного медиального контура червя мозжечка (см. рис. 1). Передняя его граница отчетливо не видна из-за гипоэхогенности дорсальной части моста. Переднезадний размер IV желудочка в неонатальном периоде не превышает 4 мм.
Мозолистое тело. Мозолистое тело (corpus callosum) на сагиттальном срезе выглядит как тонкая горизонтальная дугообразная гипоэхогенная структура (рис. 2), ограниченная сверху и снизу тонкими эхогенными полосками, являющимися результатом отражения от околомозолистой борозды (сверху) и нижней поверхности мозолистого тела. Сразу под ним располагаются два листка прозрачной перегородки, ограничивающие ее полость. На фронтальном срезе мозолистое тело выглядит тонкой узкой гипоэхогенной полоской, образующей крышу боковых желудочков.
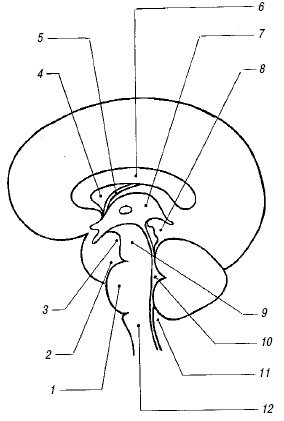
Полость прозрачной перегородки и полость Верге. Эти полости расположены непосредственно под мозолистым телом между листками прозрачной перегородки (septum pellucidum) и ограничены глией, а не эпендимой; они содержат жидкость, но не соединяются ни с желудочковой системой, ни с субарахноидальным пространством. Полость прозрачной перегородки (cavum cepti pellucidi) находится кпереди от свода мозга между передними рогами боковых желудочков, полость Верге расположена под валиком мозолистого тела между телами боковых желудочков. Иногда в норме в листках прозрачной перегородки визуализируются точки и короткие линейные сигналы, происходящие от субэпендимальных срединных вен. На корональном срезе полость прозрачной перегородки выглядит как квадратное, треугольное или трапециевидное анэхогенное пространство с основанием под мозолистым телом. Ширина полости прозрачной перегородки не превышает 10-12 мм и у недоношенных детей шире, чем у доношенных. Полость Верге, как правило, уже полости прозрачной перегородки и у доношенных детей обнаруживается редко. Указанные полости начинают облитерироваться после 6 мес гестации в дорсовентральном направлении, но точных сроков их закрытия нет, и они обе могут обнаруживаться у зрелого ребенка в возрасте 2-3 мес.
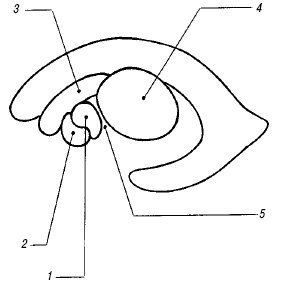
Базальная (c. suprasellar) цистерна включает в себя межножковую, c. interpeduncularis (между ножками мозга) и хиазматическую, c. chiasmatis (между перекрестом зрительных нервов и лобными долями) цистерны. Цистерна перекреста выглядит пятиугольной эхоплотной зоной, углы которой соответствуют артериям Виллизиева круга.
Ножки мозга (pedunculus cerebri), мост (pons) и продолговатый мозг (medulla oblongata) расположены продольно кпереди от мозжечка и выглядят гипоэхогенными структурами.
Паренхима. В норме отмечается различие эхогенности между корой мозга и подлежащим белым веществом. Белое вещество чуть более эхогенно, возможно, из-за относительно большего количества сосудов. В норме толщина коры не превышает нескольких миллиметров.
Стандартные эхоэнцефалографические срезы
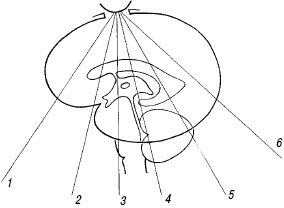
Рис. 4. Плоскости коронального сканирования (1-6).
Сосудистое сплетение головного мозга
… остается одной из наименее изученных структур головного мозга, а проблемы физиологической и патологической ликворо-динамики, представляющие существенный интерес для клиницистов-неврологов, все еще далеки от разрешения.
Среди трех частей СС наибольшее значение имеет СС конечного мозга. Оно секретирует основную массу ЦСЖ, оно чаще является местом развития патологических процессов и объектом хирургических вмешательств, в ходе его развития у человека наблюдаются наиболее значительные перестройки. Поэтому большее внимание будет уделено СС боковых желудочков мозга.
Основу СС составляют многочисленные кровеносные сосуды, которые с сопровождающими их нервными волокнами располагаются в рыхлой соединительнотканной строме, отделенной от полости желудочка одним слоем эпителия, на поверхности которого обнаруживаются клетки Колмера (см. далее). СС появляется у эмбриона человека в конце седьмой недели пренатального развития, а выработка ЦСЖ вероятнее всего начинается на девятой неделе эмбриогенеза (СС продолговатого и промежуточного мозга образуются путем инвагинации однослойной крыши, в то время как сплетения боковых желудочков [конечного мозга] развиваются из медиальной части из многоклеточного матрикса; раньше всего появляется закладка СС четвертого желудочка).
Кровоснабжение СС конечного мозга человека осуществляется при помощи одной передней хороидальной артерии, отходящей от внутренней сонной артерии, и нескольких задних хороидальных артерий, являющихся ветвями задней мозговой артерии. Отток крови обеспечивают вены основания мозга и большая вена Галена. Артерии ветвятся, образуя в конечном счете сеть широких синусоидных капилляров, имеющих локальные расширения. Имеются сведения и о существовании в СС артерио-венозных шунтов. Стенка капилляра состоит из фенестрированного эндотелия, базальной мембраны и перицитов. Количество фенестр в эндотелии тем больше, чем меньше расстояние между капилляром и эпителием сосудистого сплетения. В зоне фенестр и на люминальной поверхности эндотелиоцитов концентрируются сульфатированные протеогликаны, которые придают мембране отрицательный заряд, ограничивающий прохождение через эндотелий анионных молекул. Эндотелиоциты капилляров сосудистого сплетения содержат значительное количество белка Glut1, ответственного за транспорт глюкозы через гисто-гематические барьеры. В регуляции кровотока через сосудистое сплетение участвуют катехоламины симпатических нервных волокон, ангиотензин II и окись азота, выработку которой (как показывают экспериментальные исследования, выполненные на крысах) может обеспечивать эпителий сосудистого сплетения. Механизмы регуляции гемодинамики в сосудистом сплетении характеризуются высокой степенью автономии.
Соединительная ткань CC имеет лептоменингеальное происхождение и состоит из клеток и межклеточного вещества, которое наряду с основным веществом представлено коллагеновыми и ретикулярными волокнами. Большая часть клеток стромы имеет морфологические черты фибробластов, однако по гистогенетическим потенциям они существенно отличаются от «обычных» фибробластов соединительной ткани, расположенной за пределами органов нервной системы. Фибробласты стромы сосудистого сплетения составляют единую популяцию с менингоцитами (арахноэндотелиоцитами), которые обладают нехарактерной для типичных фибробластов способностью изменять фенотип от фибробластоподобного до эпителиоподобного. В строме сосудистого сплетения человека также присутствуют тучные клетки и макрофаги (которые имеют гематогенное происхождение).
Эпителий СС является однослойным кубическим. На апикальной поверхности эпителиоцитов имеется большое количество микроворсинок и несколько ресничек, сгруппированных в виде пучка, базальные тельца которых погружены глубоко в цитоплазму. Эпителиоциты соединены между собою плотными и щелевыми контактами, интердигитациями и десмосомами. Поверхностные клетки СС (клетки Колмера [КК]) являются макрофагами и дифференцируются из моноцитов крови, которые мигрируют на поверхность СС, проходя стенку кровеносного сосуда, соединительную ткань и эпителий. Как и тканевые макрофаги, КК имеют рецепторы к комплементу. КК являются важным компонентом гемато-ликворного барьера, так как именно они утилизируют посторонние вещества, попавшие в ЦСЖ в результате различных воздействий или несостоятельности барьерных структур. Не исключена возможность их влияния на проницаемость ГЛБ при посредстве окиси азота. Помимо выполнения защитных функций, КК участвуют в транспорте и накоплении желез. Существует мнение, что эти клетки фагоцитируют фрагменты цитоплазмы эпителиоцитов сплетения, в которых накапливаются поврежденные органеллы и включения.
Клинические аспекты патологии СС. ЦСЖ синтезируется СС бокового, третьего и четвертого желудочков. Даже небольшое нарушение баланса между продукцией и абсорбцией ЦСЖ может привести к изменению внутрижелудочкового и внутричерепного давления. Гидроцефалия чаще всего развивается в результате обструкции оттока ликвора (внутренняя гидроцефалия), либо в результате недостаточной его абсорбции в венозные синусы (наружная гидроцефалия). Иногда изменяется продукция ЦСЖ сосудистыми сплетениями. Воспаление СС или появление там папилломы может привести к гиперсекреторной гидроцефалии. Напротив, повреждение сосудистых сплетений радиацией, травмой, инфекционным агентом при менингите, либо вследствие выполнения люмбальной пункции, может привести к снижению продукции ликвора (гиполикворея). Данное состояние проявляется длительными персистирующими головными болями, которые могут исчезать и появляться при изменении положения тела.
 Подробнее о СС в следующих источниках:
Подробнее о СС в следующих источниках:
статья (обзор) «Сосудистое сплетение головного мозга и структурная организация гематоликворного барьера у человека» Д.Э. Коржевский, НИИ Экспериментальной медицины РАМН (журнал «Регионарное кровообращение и микроциркуляция» март, 2003) [читать];
статья «Морфо-функциональные особенности сосудистых сплетений желудочков головного мозга» И.Х. Гасанова, ГУ «Крымский государственный медицинский университет имени С.И. Георгиевского» (журнал «Український морфологічний альманах» №3, 2011) [читать];
статья «Морфометрические параметры капилляров ворсинок сосудистых сплетений головного мозга человека при старении» Т.М. Бабик, Кафедра анатомии Уральского государственного университета физической культуры, Челябинск (журнал «Морфологические ведомости» №1-2, 2006) [читать];
статья «Иммуногистохимическая характеристика сосудистых сплетений головного мозга человека» О.А. Юнеман, С.В. Савельев; Научно-исследовательский институт морфологии человека РАМН, Москва (журнал «Архив патологии» №5, 2012) [читать];
статья «Морфологическая организация эпифиза и сосудистого сплетения III желудочка головного мозга человека» Юнеман О.А., Лаборатория развития нервной системы ФГБУ «Научно-исследовательский институт Морфологии человека» РАМН, Москва (журнал «Морфологические ведомости» №3, 2012) [читать];
статья «Опухоли сосудистого сплетения головного мозга» Орлов Ю.А., Шаверский А.В., Свист А.А.; Отдел нейрохирургии детского возраста, Институт нейрохирургии им. акад. А.П. Ромоданова НАМН Украины, г. Киев ( Український нейрохірургічний журнал, №2, 2012) [читать];
статья «Локализация сосудистой мальфориации в сосудистом сплетении головного мозга» Молчанова А.А., Гринберг В.Б., Кушиков К.Т., Беккалиев А.Т., Лигузов Д.П.; Алматинский государственный институт усовершенствования врачей; Центр военной медицины, г. Алматы (журнал «Вестник АГИУВ» №1, 2014) [читать];
статья «Spectrum of Choroid Plexus Lesions in Children» Ramin M. Naeini, Jeong Hyun Yoo, Jill V. Hunter; Department of Radiology, Baylor College of Medicine, Houston; Dongdaemun Hospital, School of Medicine, Ewha Womans University, Seoul, Korea; Texas Children’s Hospital, Houston (American Journal of Roentgenology, January 2009) [читать] или [читать];
статья «Radiologic-Pathologic Correlation Congenital Choroid Plexus Papillomas» David R. Anderson, Steven Falcone, Jocelyn H. Bruce, Andres A. Mejidas, and M. Judith Donovan Post; From the Departments of Radiology, Pathology, and Obstetrics and Gynecology, University of Miami School of Medicine (American Society of Neuroradiology, 16, November 1995) [читать]
Какой отдел вентрикулярной системы не содержит сосудистое сплетение

Из множества аспектов исследования механизмов, обеспечивающих оптимальное функционирование центральной нервной системы, достаточно большой период времени активно изучаются структурно-функциональные возможности гематоэнцефалического барьера (ГЭБ) [1, 2].
Именно наличие ГЭБ позволяет поддерживать гомеостаз всего организма, в том числе и центральной нервной системы [3, 4]. Наиболее функционально значимой структурой признаны plexus choroids головного мозга (ССГМ) [5, 6]. Очевидно, что важным является онтогенетическое становление морфологического субстрата ССГМ [7]. Несомненно, важна морфофункциональная состоятельность ССГМ не только в условиях нормы, но с учетом окружающей среды [8, 9]. Формирование любой структуры организма хордовых, включая и млекопитающих, обусловлено также окружающей средой, в частности воздействием стресса [10].
Однако, несмотря на большое внимание исследователей к особенностям функционирования центральной нервной системы, частные вопросы структуры и функций plexus choroids оставляют достаточно много вопросов по формированию барьера, в частности, в онто- и филогенезе. Требуют детализации преобразования структурных компонентов plexus choroids в историческом аспекте. Отсутствует исследование рандомизации морфофункционального состояния plexus choroids боковых, 3-его и 4-го желудочков ликворной системы головного мозга. Совершенно очевидно, что это позволит раскрыть механизмы защиты от проникновения агрессивных препаратов, способных влиять на гомеостаз в центральной нервной системе.
Было проведено сравнительное изучение структурно-функционального становления plexus choroids головного мозга позвоночных животных боковых, 3-го и 4-го желудочков и выявление особенностей функционального созревания каждого.
В работе использованы методы: микроанатомические (Тотальный пленочный препарат сосудистых сплетений головного мозга мышей по Л.Г. Сентюровой и Р.А. Зумерову, авторское свидетельство № 1288536), гематоксилин и эозин, Ван–Гизон, Харт; толуидиновый синий, по Нахласу, по Гомори и Гленнеру; метод импрегнации по В.В. Куприянову. Проводилось определение высоты клеток хориоидного эпителия (в мкм), средний диаметр их ядер (в мкм), толщина соединительнотканной стромы (в мкм), средний диаметр отдельных звеньев микроциркуляторного русла (в мкм). Статистическую обработку полученных данных осуществляли на персональном компьютере с использованием пакета «Анализ данных» в рамках программы Microsoft Excelи BioStat 2008Professional 5.8.4.
Материалом для исследования служили ССГМ миноги (3), карповых (3) и осетровых рыб (3), земноводных (3), птиц (3) и беспородных белых крыс (25), мышей (25). Всего было детально изучены ССГМ 65 особей. При работе использовали устройство для фиксации мелких лабораторных животных в эксперименте [11].
Результаты исследования и их обсуждение
Обнаружено, что у миноги в переднем мозге plexus choroids представлены эпителиальной пластинкой. Отмечается расположение эпителия в несколько слоев. Кровеносные сосуды тесно соприкасаются с этим эпителием, что позволяет обеспечивать максимальный функциональный контакт. Сосуды заполнены элементами крови.
У карповых ССГМ кроме сплетения в желудочке продолговатого мозга, они располагаются и в промежуточном мозге. У сазана, каспийской воблы, толстолобика, plexus choroids заметно развиты лишь в 3-м и 4-м желудочках. Дупликатура видоизмененной эпендимы с одной стороны образована цилиндрическими клетками, с другой – плоской формы. Первые характеризует практически не окрашивающаяся цитоплазма и апикальное расположение ядра. Эти же взаимоотношения структур наблюдались и у взрослых особей. Вероятно, это сформировано под влиянием среды обитания и особенностями функционирования эпителиальных клеток ССГМ. А у осетра plexus choroids переднего мозга представлено дупликатурой видоизмененной эпендимы медиальной стенки желудочка. Эпителиоциты (хориоэпителиоциты) довольно однородной плоской формой. Сосуды встречаются редко. Соединительнотканная ткань не определяется. Plexus choroids 3-го и 4-го желудочков мозга были образованы сосудистыми клубочками с тонкой эндотелиальной выстилкой, обеспечивающей оптимальные условия для местного метаболизма. В то же время у севрюги мы не обнаружили plexus choroids в конечном мозге.
Достаточно позднее формирование элементов ГЭБ в классе рыб, возможно, определяется их функциональным значением. Об этом свидетельствует и тот факт, что форма их мало меняется, а только биометрические показатели.
У взрослых амфибий plexus choroids являются структурно оформленными образованиями. Эпителий, обращенный в полость желудочка, образован однослойными клетками кубической формы. Каждый эпителиоцит содержит по одному ядру, чаще округлой формы. В ядре прослеживается нежная сеточка гетерохроматина. Цитоплазма слегка базофильна. Толщина соединительнотканной стромы существенно увеличена по сравнению с филогенетически более ранними формами. В сплетениях всех желудочков наблюдается стаз элементов крови. Это указывает на существенную функциональную особенность ССГМ в процессах метаболизма у земноводных. По-видимому, медленный кровоток способствует более качественному обмену жизненно необходимыми веществами.
Сравнение plexus choroids головного мозга рептилий с земноводными позволяет утверждать, что они присутствуют также в боковых, 3-м и 4-м желудочках. Наибольшую площадь занимают сплетения в боковых и в 4-м желудочках. Строение аналогично таковому у амфибий. Хорошо выражена гроздевидная часть. Эпителиоциты преимущественно кубической формы располагается в один слой на базальной мембране. Следует отметить, что высота эпителиоцитов сравнима с аналогичными клетками амфибий. Каждая клетка содержит по одному ядру округлой формы и содержит довольно много гетерохроматина. Под базальной мембраной располагается рыхлая соединительная ткань. В её толще залегают довольно многочисленные кровеносные сосуды. Присутствуют фибробласты, наблюдаются единичные пигментоциты. Меньше всего соединительной ткани в сосудистом сплетении третьего желудочка.
Впервые у птиц закладку plexus choroids сосудистых сплетений головного мозга можно наблюдать на 2-й день пренатального развития. Первыми образуются сплетения 4-го желудочка в виде складки эпендимы с зародышевой мезенхимой. Только к концу первой недели начинается гистогенез plexus choroids боковых желудочков. Однако, эпителий псевдомногорядный. Ближе к вылуплению он становится однорядным однослойным. Как и у более ранних эволюционных форм каждый эпителиоцит (хориоэпителиоцит) имеет ядро чаще овальной формы. Можно наблюдать ядрышко и небольшое количество гетерохроматина. Цитоплазма оксифильна. Лишь к середине второй недели нами наблюдались plexus choroids 3-го желудочка. После вылупления в plexus choroids регистрировалось продолжение морфофункциональной дифференцировки, при сохранении заметной гетерохронии созревания ССГМ боковых, 3-го и 4-го желудочков.
В классе млекопитающих (крысы, мыши) сначала формируются ССГМ 4-го желудочка. В начале 3-й недели (15–17 день пренатального развития) появляются закладки ССГМ в боковых желудочках и 3-го желудочка. Далее гистогенез сосудистых сплетений продолжается уже в постнатальном периоде, когда в 3-м и 4-м желудочках появляются ворсинки, увеличивающие поверхность для оптимального обмена веществ. На пленочных препаратах показано, что формирование сосудистого русла продолжается вплоть до полного созревания организма. Сравнительный анализ структуры ССГМ желудочков позволяет сделать вывод, что наибольшую функциональную нагрузку несут ССГМ третьего и четвертого желудочков.
ССГМ у человека имеют значительно большие размеры и сложность строения. В 3-м и 4-м желудочках plexus choroids образовано инвагинацией однослойной крыши. А plexus choroids боковых желудочков являются производными медиальной части матрикса. Раньше других наблюдается закладка сосудистого сплетения 4-го желудочка. Очень рано ССГМ начинают формировать вторичные ворсины. В 7 недель эмбрионального развития сплетения боковых желудочков имеют лишь короткие первичные ворсины. И только на 8-й неделе появляются первые ветвления. Строма построена из мезенхимы, содержащей различные клеточные элементы. В плодном периоде к 15 неделям plexus choroids заполняют большую часть полости боковых желудочков. В этот период ядра эпителиоцитов (хориоэпителиоцитов) располагаются ближе к апикальному полюсу, что говорит о высокой степени метаболических процессов, в частности секреторных. После 16 недель ядра эпителиоцитов занимают обычное положение (ближе к основанию клетки). Наличие гломуса в сосудистых сплетениях бокового и 4-го желудочков, оставляет необходимость дальнейшего изучения plexus choroids, для более полного представления о роли ССГМ в обеспечении гомеостаза центральной нервной системы.
Сравнительное исследование филогенеза и отдельных этапов индивидуального развития сосудистых сплетений головного мозга (plexus choroids) миноги, рыб, земноводных, птиц, млекопитающих и человека позволило выявить этапы их морфогенеза и структурно-функционального становления гематоэнцефалического барьера.
Так, у хордовых основными закономерностями морфогенеза сосудистых сплетений головного мозга можно считать следующее: первичные сосудистые сплетения миноги являются простейшими образованиями и построены из эпителия. Последний представлен несколькими слоями. Сосуды лишь прилежат к нему.
У рыб начинают формироваться однослойные эпителиальные тяжи, окаймляющие примитивную соединительную ткань. Эпителиоциты резко не равноценны по высоте.
У амфибий ССГМ присутствуют во всех желудочках: боковых, третьем и четвертом. У них формируются не только первичные, но и вторичные ворсины. При этом сохранен план строения: однослойный эпителий расположен на базальной мембране. Клетки кубической формы, иногда уплощенные. В первых ядра округлые, во вторых – овальные. Все они содержат по 1–2 ядрышка. В соединительной ткани множественные сосуды микроциркуляторного русла. Наблюдаются единичные фибробласты.
У рептилий наибольшую функциональную нагрузку несут сосудистые сплетения боковых и четвертого желудочков. Именно они наиболее мощно представлены структурно. В этом классе хордовых эпителий представлен аналогично по сравнению с более низшими эволюционными формами. Но соединительнотканная прослойка содержит большее количество клеточных элементов, среди которых фибробласты, единичные фиброциты и пигментоциты.
У птиц закладка сосудистого сплетения формируется в боковых желудочках в виде многослойного эпителия с дальнейшей реорганизацией в однослойный. Порядок сходен с таковым у млекопитающих. Также наблюдается схожее строение в силу множественного ветвления ворсин в боковых желудочках головного мозга как у человека. Эпителий сосудистых сплетений однослойный кубический. Расположен на базальной мембране. Соединительнотканная прослойка представлена рыхлой соединительной тканью с сосудами микроциркуляторного русла и клеточными элементами.
Анализируя полученные морфометрические данные на примере ССГМ млекопитающих – мышей (рис. 1), можно отметить, что высота эпителиоцитов plexus choroids (хориоэпителиоцитов) 4-го желудочка во все сроки развития по размерам превышает размеры эпителиоцитов в боковом и 3-м желудочке (p 0,05).
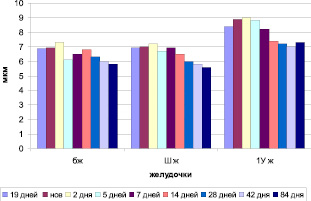
Рис. 1. Морфометрия сосудистых сплетений головного мозга мышей в онтогенезе (эпителий)
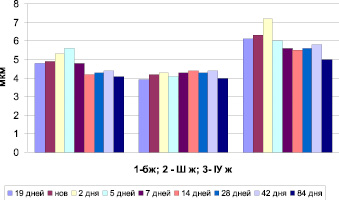
Рис. 2. Морфометрия сосудистых сплетений головного мозга мышей в онтогенезе (ядра)
У новорожденных мышей только в четвертом желудочке наблюдается увеличение высоты эпителия (p 0,05), хотя эпителий имеет тенденцию к снижению высоты (p 0,05).
Заключение
Исследование plexus choroids головного мозга позвоночных в сравнительном аспекте позволило установить ряд закономерностей, свидетельствующих об особенностях становления морфофункционального субстрата гематоэнцефалического барьера. Процессы дифференцировки наблюдаются во всех основных морфологических структурах, в частности в хориоэпителиоцитах.